现代丝绸之路东段植物物种多样性格局及其影响因素研究

iMeta主页:http://www.imeta.science
研究论文
● 原文链接DOI: https://doi.org/10.1002/imt2.74
● 2023年1月9日,中国科学院植物研究所周世良团队与河北工程大学刘艳磊团队在iMeta 在线发表了题为"What determines plant species diversity along the Modern Silk Road in the east?"的文章。
● 本研究沿着中国西北荒漠区现代丝绸之路东段进行了植物物种多样性调查,发现在这一区域温度比降水更能影响当地的植物物种多样性。此外,人类活动可能通过引进作物和入侵植物以及消除适应环境的本地植物来改变当地的植物物种多样性格局。研究证明了基于eDNA的宏DNA条形码技术在植物物种多样性调查方面的潜力。
● 第一作者:刘艳磊
● 通讯作者:周世良(slzhou@ibcas.ac.cn)、王宇飞(wangyf@ibcas.ac.cn)
● 合作作者:徐超、董文攀、陈荀、张稳、孙雨哲、王国宏
● 主要单位:
河北工程大学园林与生态工程学院、中国科学院植物研究所系统与进化植物学国家重点实验室、中国科学院大学生命科学学院、北京林业大学、呼伦贝尔学院、中国科学院植物研究所植被与环境变化国家重点实验室
亮 点

● 环境DNA可以快速高效评估植物物种多样性
● 首次利用环境样品DNA(eDNA)评估大尺度区域性的植物物种多样性格局
● 温度而不是降水是影响当地植物物种多样性的主要因素
● 人类活动影响了荒漠区当地的植物物种多样性
● 治理荒漠化需要取得更多更及时的植物多样性结果
摘 要
作为生产者,植物为全球生态系统提供了食物、氧气和其他资源,在生物多样性保护中理应得到优先的保护。目前,多数研究集中在生物多样性热点区域,诸如荒漠区的生物多样性冷点区域大部分被研究者遗忘。我们认为影响生物多样性热点和冷点区域的主要因素可能存在很大的差异,尤其是对于荒漠生态系统而言。为了验证这一猜想,我们沿着中国西北荒漠区现代丝绸之路东段进行了植物物种多样性调查,这一区域有降水量低、植被稀少、物种数量有限、不同区域人类活动差异大的典型特征。本研究在该区域采集得到144份表土样品,通过土壤中的环境DNA(eDNA)和宏DNA条形码技术(Metabarcoding)来揭示当地的种子植物物种现状。经过分析,本研究共发现种子植物671种,这一结果远高于我们在地上植物调查的结果。同时,本研究发现在这一区域温度比降水更能影响当地的植物物种多样性。此外,人类活动可能通过引进作物和入侵植物以及消除适应环境的本地植物来改变当地的植物物种多样性格局。我们的研究证明了基于eDNA的宏DNA条形码技术在植物物种多样性调查方面的潜力。荒漠植物经过长期演化,已经演化出利用地下水或利用偶尔的降雨作为短暂水源的干旱环境适应机制,由于他们地上部分的生命周期短,这些植物在地表植物调查时很难被发现和鉴定,但基于eDNA的宏DNA条形码技术帮助我们挽回了这一损失。最后,我们建议在当地植物物种多样性保护和荒漠化治理中将重点放在地下水的维持和人类活动的控制当中。
视频解读
Bilibili:https://www.bilibili.com/video/BV1dG4y1k78w/
Youtube:https://youtu.be/Xcty5Z8UYUw
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
生物多样性是衡量地球活力的指标,也是人类生存和发展的基础。因此,保护生物多样性便是保护人类自己。作为初级生产者,植物为全球生态系统提供了食物、氧气和其他资源,理应在生物多样性保护当中给予优先重视。获得植物物种多样性状况的传统方法仍然是地表植物调查,这类方法需要多年大量的样品采集和实地踏查,资金投入大、耗时长、有很高的劳动力要求。此外,在当今植物分类学家逐渐濒危的时期,标本的准确鉴定渐渐地成为了一种挑战,一些体型较小或微小的植物在调查过程中很有可能被忽略。最近出现的宏DNA条形码:一种基于环境DNA的研究方法使得快速鉴定环境样品中的植物信息成为可能。eDNA是从特定环境样品中提取的DNA,这些环境样品可以是土壤、水、粪便或者空气,这些样品中可能包含了生活在其周边区域的植物、动物和微生物的残体或游离DNA。宏DNA条形码是一类基于高通量测序的宏基因组学技术,不同物种的混合DNA可以被同时测序,测序结果可以利用DNA条形码技术进行物种鉴定。
基于环境样品的宏DNA条形码已经被应用于古生物多样性重建、现代生物多样性评估、生物监测、入侵物种评估、动物食性评估、植物与微生物互作和物证鉴定分析等领域。尽管有一些综述类文章提及了利用环境样品进行植物多样性研究的可能性,但基于eDNA的大尺度植物物种多样性研究仍未出现。由于植物的记录情况远优于动物和微生物,因此,植物是基于环境DNA的理想研究对象。中国已经相继完成了《中国植物志》和《Flora of China》, 中国植物DNA条形码参考数据库也已经建成。因此,摆在我们面前唯一的挑战便是代表性土壤样品的采集。当研究当代的植物物种多样性时,采集不深于5厘米的表层土壤样品可以反映近几十年的植物信息。前人的研究主要是关于动物和微生物的,这二者对于样品采集的策略与植物略有不同。不同于植物,动物的活动范围较大,因此需要对土壤样品进行深层取样。对于微生物,他们的拷贝数量十分庞大,这使得采集少量土壤便可以得到大量的微生物信息,因此,采样深度不会限制微生物相关的研究。植物将他们的DNA以游离DNA或者DNA载体的形式保存在他们生活的年代地层中。通常来讲,地层的深度越大,反映的植物信息越古老。尽管基于表层土壤样品的植物物种多样性研究很少,前人的研究仍然告诉我们表层土壤样品中的eDNA可以反映现代植物的信息,因为从土壤中获得的植物物种的多样性比在地面上生长的植物物种更丰富。
多数关于物种多样性的研究聚焦在生物多样性热点区域。荒漠区以降水稀少、土壤贫瘠、生态环境恶劣、生多样性低为特征,是大众熟知的生物多样性冷点区域。此类生物多样性冷点区域通常植被稀少,对周边环境变化和人类活动较为敏感。当今世界正在经历因大面积荒漠化带来的生物多样性冷点区域扩张和生物多样性热点区域收缩。作为世界上第二大荒漠区--亚欧荒漠从中亚地区一直延伸到地中海东海岸,与丝绸之路的路径重合。中国西北荒漠区坐落在丝绸之路东段,这一区域见证了丝绸之路沿线诸多文明的兴衰,曾经在东西方文化交流中扮演重要的枢纽作用。前人在此区域的群落调查研究揭示了当地贫瘠的生物多样性和以小型短生命周期草本植物和抗旱草本植物、灌木和小乔木为特征的植物群落。在本研究中,我们将利用宏DNA条形码技术探索中国西北荒漠区的植物物种状况,验证以下假设:(1) 环境样品反映的植物物种多样性高于植物群落调查显示的多样性; (2) 影响生物多样性热点和冷点区域形成地理植物多样性模式的因素不同。荒漠区常有强风,可以相对均匀地散布植物残体,因此,地表土壤和植物凋落物的收集比植物群落调查更能代表当地植物区系。刚完成地上生命周期或从某个地区消失的物种很可能会在表层土壤中沉积可检测到的植物残骸。此外,在某些器官缺乏物种鉴定特征的情况下,例如在植物区系相对简单的荒漠中,通过宏DNA条形码进行物种识别更为容易和准确。荒漠植物群是演化和适应当地环境因素(如降水、温度、土壤和人类活动)的结果,造成当地生物多样性丧失的主要因素以及如何有效保护荒漠植物物种多样性仍有待探索。因此,我们采集了中国西北荒漠区丝绸之路东段144份表层土壤样本并记录了周边的气象记录和人口状况,主要回答以下问题:(1) 这一区域种子植物物种多样性的空间格局如何? ; (2) 哪些因素在当地植物物种多样性空间格局的形成中贡献最大?;(3) 人类活动如何影响当地植物物种多样性? 本研究的结果将提高研究者对中国西北荒漠区植物物种多样性的认识,并有助于制定保护生物多样性、治理荒漠化和恢复荒漠植被的战略政策。
结果
DNA条形码参考数据库
DNA条形码参考库由三个数据集组成:rbcL 、matK 和trnL -intron。rbcL数据集包含80科361属853个种子植物物种,matK数据集包含81科374属944个种子植物物种,trnL-intron包含70科316属686个种子植物物种。三个DNA条形码参考库共包含83科432属1,183种,占该地区物种的76%、属的86%和科的95%。
土壤样品中发现的种子植物
最终,rbcL , matK 和trnL -intron从Illumina 测序平台分别获得了3.02, 2.29和19.08个百万的clean reads. 将这些clean reads mapping 到相应的参考数据库,最终144个表层土壤样品共计发现65科,291属,671个植物物种 (图2)。其中,涵盖62科,266属,623种当地野生植物物种;20种入侵植物;28种栽培植物。从reads相对丰富度的角度来看,丰度最高的5个科分别为Asteraceae (16.11%), Rosaceae (12.50%), Pinaceae (6.73%), Nitrariaceae (6.09%), 和 Fabaceae (5.94%);丰度最高的5个属分别为Artemisia (7.73%), Picea (6.68%), Nitraria (5.87%), Juniperus (4.36%) 和Ulmus (4.34%);丰度最高的10个物种分别为Nitraria sibirica (2.63%), Iljinia regelii (2.30%), Ulmus glaucescens (2.17%), Ulmus pumila (2.17%), Picea asperata (1.67%), Picea crassifolia (1.67%), Picea obovate (1.67%), Picea schrenkiana (1.67%), Typha angustifolia (1.29%) 和 Morus nigra (1.28 %)。从物种数量上来看,物种数量最多的5个科分别为 Poaceae (14.13%), Asteraceae (12.20%), Rosaceae (10.91%), Fabaceae (7.06%) 和 Ranunculaceae (5.62%)。
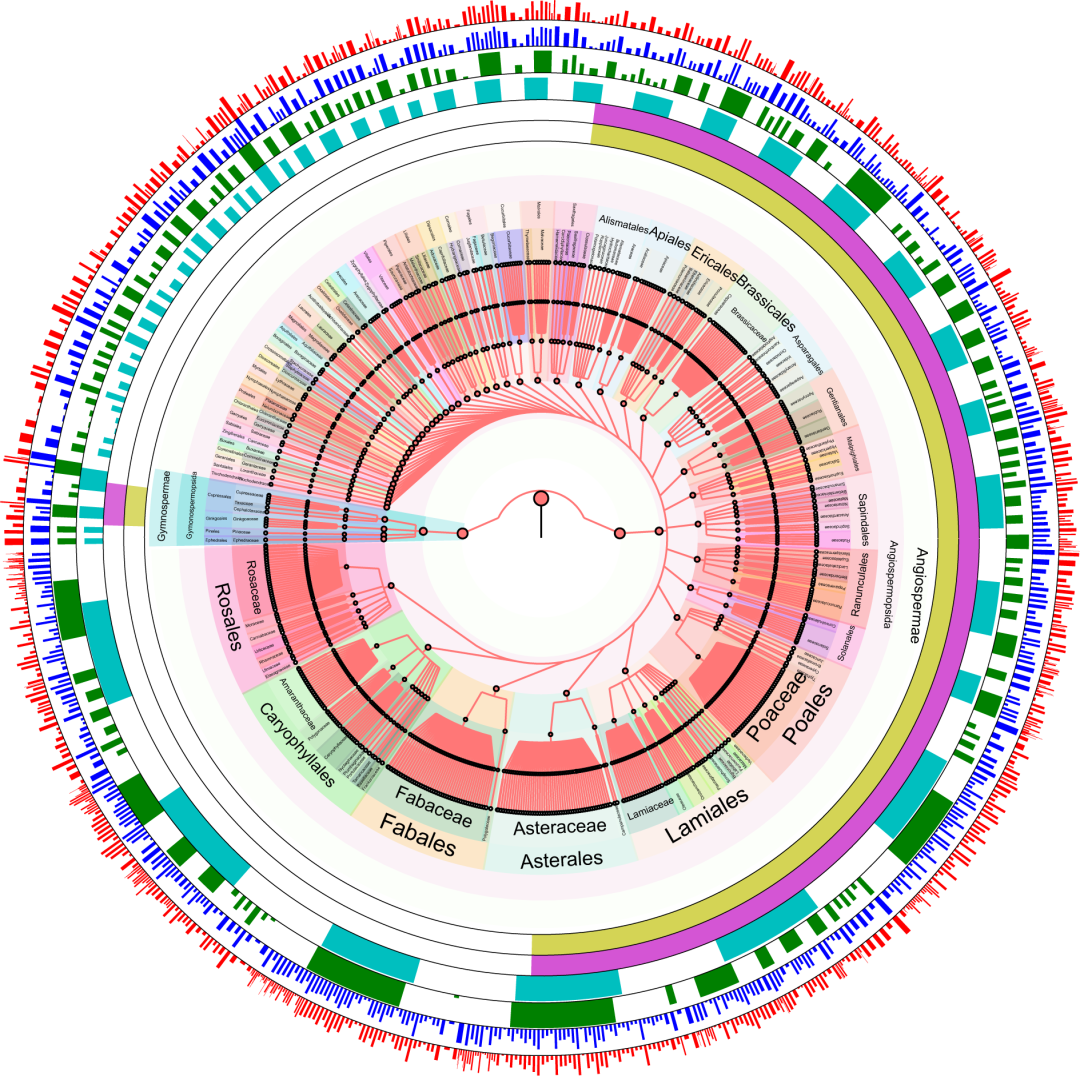
图2. 基于土壤表层样品的中国西北荒漠区种子植物整体情况
地表样方调查中的种子植物
通过地表样方植物调查,共计发现23科,59属,共计87个植物物种。Amaranthaceae (20 species), Fabaceae (13 species), Asteraceae (13 species), Poaceae (8 species) 和 Zygophyllaceae (6 species) 是其中物种最多的五个科,Artemisia (5 species), Caragana (5 species), Salsola (5 species) 和Zygophyllum (4 species) 是其中物种数量最多的属。在发现的这些物种中,44个物种也同时在环境样品中被发现,占样方调查结果的50.6%。
植物物种多样性格局
利用每个采样点的种子植物组成,包括本地野生、栽培和入侵物种,获得了植物物种多样性的地理分布格局。本区域中部地区的植物种类数量和相对丰富度比两侧外部地区复杂。中部地区的植物物种组成处于平衡状态,没有明显的优势物种(图3A)。例如,040号样点有383种植物,其中葡萄的比例最大,为3.14%。相比之下,Enneapogon desvauxii 在东部样点012处显示出29.46%的优势,而Iljinia regelii在西部样点137处显示出25.68%的优势。Shannon.e多样性指数显示了本地野生物种(图3B)、栽培物种和入侵物种(图3C和D)的相似趋势。中国西北荒漠区植物物种多样性通常从东到西逐渐减少,这一趋势的最大例外是东部的20个地点,这些样点主要分布在乌兰布和腾格里沙漠,由于原本几乎不含任何植被,这些地点的植物物种多样性较低。去除这些样品后,中国西北荒漠区呈现出东高西低的植物物种多样性分布格局。
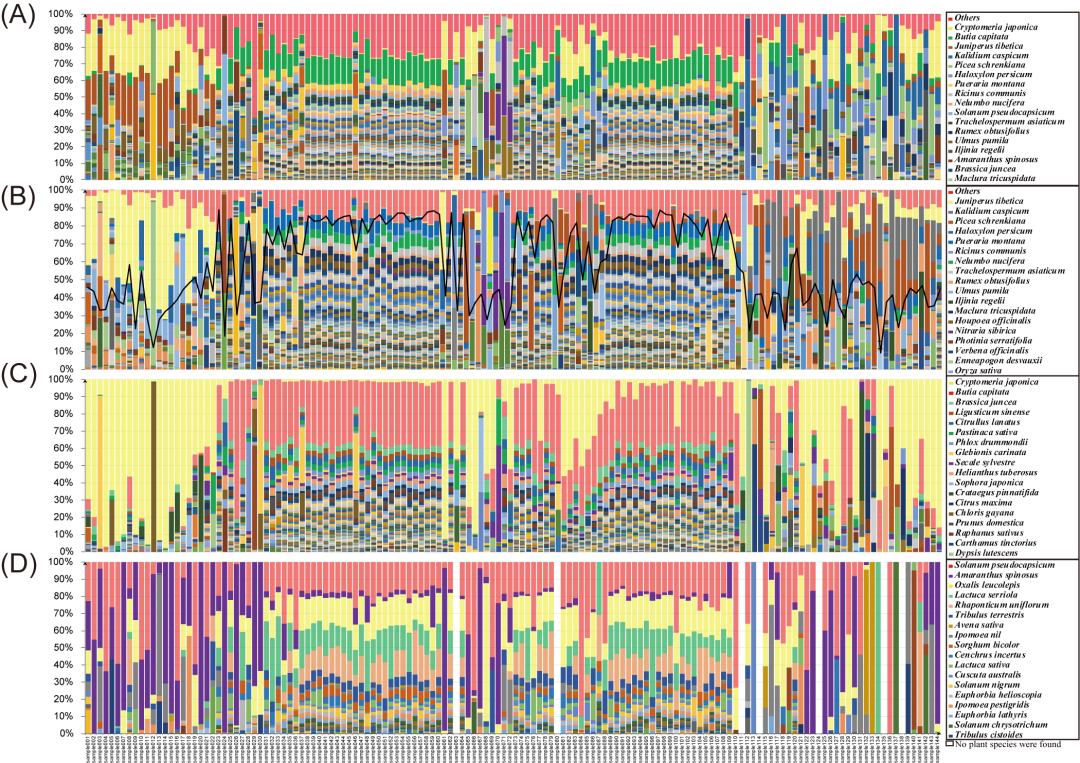
图3 基于各样点表土中植物物种组成的中国西北荒漠区植物物种多样性分布格局
(A): 代表所有种子植物; (B): 代表所有野生种子植物; (C): 代表所有栽培种子植物; (D): 代表所有入侵种子植物。
植物物种多样性与生态因素的关系
分析结果表明,Shannon.e多样性指数与海拔和经度呈正相关(P < 0.05 ), 与近30年的年均温、1-12月均温、最暖月最高温、最冷月月均温、最冷季均温、最暖季均温、最暖季均温和无霜期呈负相关 ( 图4 & 5)。Chao1 多样性指数与海拔高度、经度和各季节均降水呈正相关(P < 0.05 ),与温度相关因素呈负相关(P < 0.05 ) (图5)。其他α多样性指数和因子之间的相关性表现出与Shannon.e多样性指数和Chao1多样性指数相似的趋势。

图4. Shannon.e多样性指数与潜在影响因素的关联分析。
Shannon.e多样性指数与(A): 近30年均降水; (B): 地表物种数量; (C): 人口密度; (D): 最冷季均温; (E): 经度; (F): 海拔高度。

图5. 各类多样性指数与潜在影响因素的关联分析结果
不同颜色的线条用于表示相关性分析的结果是否具有统计学意义。
植物物种多样性与人口密度的关系
人口密度是人类活动对环境干扰的一种体现。分析发现人口密度与本地的植物物种多样性呈现显著负相关 (图5)。UPGMA树状图显示了植物物种多样性与人口密度之间的关联,在人类聚居区较多的地区,本地植物物种多样度较低(图6)。例如样点023的Shannon.e多样性指数最大为4.61(图6中的紫色圆圈),而样点134的Shannon.e多样性系数最小为0.359(图6的绿色圆圈)。这些结果表明,人类活动与西北荒漠区植物物种多样性的下降有关。

图6. 基于荒漠地区植物物种多样性的UPGMA聚类图
基于每个采样点野生植物物种组成的UPGMA聚类,获得了分支图和植物组成热图。地图上标出了植物物种多样性状况和人类主要聚居地(红点代表植物生物多样性丰富的区域;蓝点代表植物多样性稀有区域;红色圆圈加加号的符号代表了人类主要聚居地)。
讨 论
基于环境样品的宏DNA条形码技术是一种快速、准确且经济高效的植物多样性研究方法
地表植物调查以往是评估植物物种多样性的唯一可行方法,但费用昂贵、耗时且劳动密集。现在,基于环境样本的宏DNA条形码成为一种可行的替代方法。本研究与同一地区传统样方调查结果的直接比较得出了基于更可靠数据的植物物种组成、当地优势物种和物种多样性地理模式相同的结论 (图3)。
本研究与地表植物调查研究结果之间的一个显著差异是检测到的物种数量:分别为671种和87种。鉴于采样面积和现有物种多样性,我们认为本研究中检测到的物种数量更为合理。植物器官,如花粉粒,可以通过风或水流从其分布范围外携带到样地中,并且可以通过eDNA和DNA条形码技术相结合来检测。从样点中出现和消失的短命植物可通过eDNA宏DNA条形码根据其残留进行识别。尽管基于环境DNA的植物宏DNA条形码技术很强大,但我们的方法仅鉴定了传统样方调查中观察到的植物物种数量的约一半(50.6%)。迄今为止,类似的研究很少进行,与我们的研究不同,目前的大多数研究都集中于某些类群,并基于该类群构建参考数据库。在土壤研磨之前,我们从样品中去除了比较大的植物残体,这样可以减少获得的DNA中植物之间的偏差,我们推测这可能是造成这种差异的原因之一。此外,我们的研究比较了地上和环境样本中的植物多样性,发现这两种多样性之间没有直接的相关性。地表植物代表了取样当年的植物物种多样性,而环境样本代表了近几十年的植物种类多样性,因此直接比较从两种方法得到的结果是不合理的。在地表发现的物种数量少且随机,这也不同于进行大规模和长期地表植物调查的传统方法,因此,仅基于一年的地表植物信息统计不能反映该地区植物物种多样性的现状。此外,在eDNA宏DNA条形码缺失的43种植物物种中,19种属于苋科(原藜科),7种属于菊科,5种属于豆科。当使用传统的叶绿体片段时,DNA条形码对系统发育关系较近的物种具有低分辨率的缺点,因此在数据处理期间,系统发育关系较近的未检测到的物种的序列可能被视为背景噪声舍弃了。核糖体RNA基因的大小亚基之间的转录间隔区(ITS)可以很好地用于物种识别,但eDNA通常包含大量微生物DNA,植物ITS序列可能被微生物的ITS序列淹没,不能拿到足够的有效数据。
与传统的样方调查方法相比,eDNA宏DNA条形码技术具有以下几个优点:易于使用、速度快、节省劳动力、采样时间不受限制,并且不需要具备扎实的形态分类学背景知识。基于eDNA的宏DNA条形码技术在短时间内动态监测大尺度地理范围内的植物物种多样性方面潜力巨大。标签PCR技术的逐渐成熟也使得大量样品的同时扩增和测序成为现实。使用标签PCR标记的扩增子序列实现了对多个样本进行混合测序,这一技术将节省原始NGS文库构建、文库检查和冗余测序序列的成本,将成本降至原来的约十分之一。研究者利用有限的资金便可以获得大量扩增子数据,将有助于为这一领域的研究注入新的活力。
生态因子对植物物种多样性的影响
根据环境控制论,气候、物理和其他生态因素是多尺度植物物种多样性模式形成的主导因素。水和能量(主要是温度)是影响植物多样性的两个最重要的环境因素。在本研究中,降水与植物物种多样性没有关联性,与潮湿的热带和亚热带地区不同,荒漠地区的降水太少,无法在植物物种多样性的形成中发挥重要作用,地下水可能成为荒漠植物主要依赖。与此相反,取样地区的年平均气温从西北向东南呈下降趋势。中国西北荒漠区的植物物种多样性与温度呈负相关,前人根据传统调查数据首次观察到这一现象。这一趋势的一个可能机制是:较高的温度导致更多的水分蒸发,从而加剧干旱并降低当地的植物物种多样性。
植物所处纬度、经度和海拔是能源、水(尤其是地下水)和土壤等因素的综合体现。在本研究中,纬度与植物物种多样性没有显著相关性,这可能是因为采样点之间的纬度范围较小(标准偏差=1.2°)。虽然降水的影响似乎无关紧要,但经度与植物物种的多样性呈显著正相关。由于西部海拔较高,水(尤其是来自高山冰川融化形成的地下水)向东流,塔里木河(中国最长的内陆河流)便是这样形成的,东部地区享有更多的地下水供应,得以保持较高的植物物种多样性。由于物种及其分布均匀性的减少,植物物种多样性随着海拔的增加而减少,证实了Wang等的发现。为了了解哪些因素对植物物种多样性地理格局的形成贡献最大,我们基于Shannnon.e多样性指数对26个潜在影响因素进行了PCA分析。结果显示前16个影响因素与温度有关,表明温度在荒漠地区的重要性,经度是第二大影响因素。
人类活动影响植物物种多样性
人口的增长以及人类活动范围和强度的增加导致了植物栖息地的丧失和割裂,严重威胁到植物物种的生存和繁殖。人口密度沿着采样带从东到西逐渐增加,与植物物种多样性呈负相关(图4和5)。在PCA结果中,人类活动是影响植物物种多样性的第三大因素,表明人类活动对当地物种多样性产生了负面影响。鉴于栖息地恢复在对环境变化敏感和生态系统相对脆弱的荒漠生态系统中更加困难,限制人类活动,如过度开发和农业活动,可能是保护荒漠生态系统并保持其生态真实性的最简单方法。
荒漠植物物种多样性保护
荒漠干燥,夏季炎热,冬季寒冷,荒漠植物在这些有限的条件下适应栖息地。在中国西北荒漠区,有一些管理良好的旱生植物,如Zygophyllum spp., Haloxylon spp. 和 Calligonum spp.;深根植物,如Atriplex patens (Litv.) Iljin, Alhagi sparsifolia Shap. 和 Tamarix chinensis Lour.;短生命周期植物,如Erodium oxyrrhynchum M. Bieb., Alyssum liniolium Steph . ex Willd. 和 Schismus arabicus Nees。然而,寄生类植物,如锁阳、肉苁蓉主要依靠树木或灌木。许多荒漠植物物种都是当地特有的,如Ammopiptanthus mongolicus (Maxim. ex Kom.) Cheng f., Potaninia mongolica Maxim., Pugionium spp.、Stilpnolepis centiflora (Maxim.) Krasch.、S. intricata (Franchet) C. Shih.、Tetraena mongolica Maxim. 和 Tugarinovia mongolica Iljin。由于喜马拉雅山脉的隆起,它们被认为是第三纪古地中海的孑遗植物类群。许多荒漠特有植物是具有高经济价值的草药,例如,紫草:用于治疗麻疹、便秘、烧伤、冻伤、湿疹和皮炎,而锁阳和肉苁蓉用作壮阳药。由于过度开发,许多具有经济价值的荒漠植物已被列为濒危物种。
与生物多样性热点相比,荒漠植物物种多样性低,植被覆盖率低。地面植被需要很长时间才能建立,并且可能会瞬间遭到破坏。物种多样性与人口密度之间的负相关结果表明,西北荒漠区的人类活动导致了荒漠植物物种多样性的丧失(图4和图5)。此外,人口稠密的地区经历了更多的入侵物种和本地物种的失衡,需要迫切保护植被数量和植物物种数量。
eDNA和宏DNA条形码技术对于监测本地植物物种多样性变化和入侵植物的动态变化应该非常有效,这对有效保护当地的生态环境至关重要。宏DNA条形码技术已成功应用于微生物和动物以及本研究中的植物,我们的研究结果将通过快速准确地评估大尺度植物物种多样性、建立国家荒漠公园或自然保护区以及驯化具有经济价值的物种来促进当地的植物物种多样性保护。在大部分西北荒漠区限制人类活动似乎不太现实。与之相对应的,建立少数具有高度荒漠植物物种多样性的被保护的国家公园或自然保护区可以合理管理人类活动,并引入其他濒危荒漠植物进行保护。通过驯化具有经济价值的物种,如紫草,允许耕种并防止从脆弱的生态系统中觅食,将有助于当地的植物物种多样性保护。作为寄生植物,锁阳和肉苁蓉是更困难的驯化例子,但仍有潜力。
结 论
基于从丝绸之路东段沿线采集的144份中国西北荒漠区表层土壤样本中提取的eDNA识别的植物物种,本研究揭示了当地的植物物种多样性空间格局,共检测到671种植物,表明植物物种多样性并不像地表植物群落调查所表明的那样低。分布在该区域的优势物种主要来自菊科、蔷薇科、禾本科、松科、白刺科和豆科。植物物种多样性东部高于西部,受温度影响大于降水影响。地下水对荒漠植物的生存似乎更为重要,因此,维持高地下水位对防止荒漠化至关重要。人类活动通过减少本地野生物种,引入作物和入侵植物,对本地的植物物种多样性造成了负面影响。我们建议使用eDNA宏条形码技术对荒漠植物物种多样性进行动态监测,掌握荒漠区植物物种多样性的变化趋势。驯化经济植物是阻止当地人过度开发自然资源的一个可能的解决方案。
方 法
研究区域及土壤样品采集
本研究的样品采集时间为2015年6月8日-7月21日,采样带全长约3,600千米,经度跨度超过30 ° (E 76.313 ° 到E 106.319 ° 之间),纬度跨越超过5 ° (N 37.266 ° 到N 43.390 ° 之间),海拔高度跨度超过1,800米 (海拔239 m到海拔2,066 m之间) (图1)。取样地涵盖三类荒漠植被带,包括温带灌木草原荒漠植被带,温带灌木、半灌木荒漠植被带以及暖温带灌木、半灌木荒漠植被带。环境因子在样点间变化明显,平均年均降水量变化在221 mm (内蒙古) 到30 mm (塔里木盆地中部)之间,几乎有一半的降水集中在夏季。年均温跨度在5.2 ℃ 到 12.4 ℃之间,夏季月均温在18.1 ℃ 到28.5 ℃之间,冬季月均温在 -10.7 ℃ 到 -2.5 ℃之间,人口密度在0.166 到 84.707 人/km2之间。根据Species2000 和 GBIF记载,当地包含87科504属共计1,559种种子植物,这些植物信息将作为宏DNA条形码数据分析中的上限。

图1. 中国西北荒漠地区丝绸之路东段表层土壤样本采集点
根据采样点的位置信息使用红点标记采样点。
本研究从内蒙古西南部(阿拉善地区)沿河西走廊和新疆塔里木盆地共计获得表层土样144份。表土取样采用了经典的五点取样法,每个样方 (1,000 cm×1,000 cm) 采集5份表土样品后等量均匀混为一份,样品经65 ℃干燥24小时防止DNA的进一步降解,最后放在-20 C直至DNA提取。
生态数据收集
土壤样本中的植物物种多样性是近几十年来植物物种多样的随机样本。因此,最近30年的平均值被用作气候指标。每个样方的气候指标包括过去30年的平均年温度、月均温、季节性均温、年降水量、月降水量和季节性降水量,这些数据均从WorldClim (http://www.worldclim.org) 根据每个采样点的位置信息进行调取。采样点所属城市人口密度、主要人类聚居地(乡镇以上,包括乡镇)和干燥度(年蒸发量和干旱指数)也从每个相关城镇的网站上收集。地表土壤采样时记录了地表植物种类,仅代表当年的植物多样性。
土壤DNA提取
在进行正式的DNA提取之前,我们比较了常用的PowerSoil 土壤DNA提取试剂盒(Thermo Fisher Scientific,USA)和传统的mCTAB植物DNA提取法。结果显示二者均获得了主带明显的土壤eDNA,在rbcL1片段扩增时的表现也一致。鉴于该试剂盒的回收效率满足需求并节省时间,在对DNA提取进行了几次试点研究后,我们最终选择了PowerSoil DNA提取试剂盒用于本研究中的144个表层土壤样本的DNA提取。在进行DNA提取前,使用研钵和杵将10 g 每个样品充分混合的土壤磨成细粉。我们按照试剂盒制提供的DNA提取说明对每个样品进行DNA提取。每个样品进行三次DNA提取重复以降低物种随机偏差的可能性,并将来自同一样品的三份所得DNA合并。使用不含DNA的超纯水作为阴性对照,以检测潜在的实验室DNA污染。
PCR扩增
考虑到公共数据库的物种相对覆盖率,rbcL, matK 和 trnL-intron 这三个植物特有的叶绿体片段被采用作为本研究的DNA条形码。为了适应Illumina测序平台的读长要求,我们采用了中间引物保证rbcL 和matK的扩增片段在400 bp左右。用于扩增rbcL的引物为rbcLbF 和rbcL717LR;用于扩增matK的引物为matK472F 和 matK821R;用于扩增trnL-intron的引物为trnLc 和 trnLh。根据前人的扩增子测序方法利用Illumina测序平台进行高通量数据获取。本研究设计了24对8碱基核苷酸标记,每对引物的5'端都增加了8碱基的核苷酸标记用于区分同一高通量测序文库中的不同样品。PCR的流程参照Dong等的研究,为了减少PCR扩增中的偏好,每个相同样品PCR都进行了三次重复并将这三次重复结果合并后用于后续操作。PCR产物利用1.5%的琼脂糖凝胶电泳进行检测
DNA条形码参考数据库构建
为了更为准确的将各DNA条形码的序列鉴定至物种,我们从中国植物DNA条形码参考数据库获得了rbcL, matK 和 trnL-intron的所有分布在中国西北地区的种子植物数据,成功地构建了这三个DNA条形码片段的本地数据库。
高通量数据处理
数据处理的方法和工具参考前人的研究,原始数据的质量控制采用了NGS QC toolkit v2.3.3 (默认参数),正反向序列对接利用Pandaseq v2.11默认参数进行,matK 和 rbcL文库中相同数据库中的不同基因采用fqgrep v0.4.4 根据引物的差异进行数据拆分,拆分后的单barcode数据利用FASTX v0.0.13 toolkit将数据根据8碱基标记拆分至样品。
数据分析
- Feature table 准备
Feature table 是对每个样品中各种植物的原始序列(reads)的总结。我们利用Usearch v10.0.240, 采用0.99相似性将原始数据mapping到相应的参考数据库上从而构建相应的feature table。由于本研究采用了三个不同的叶绿体基因组片段,相应的便产生了三个feature table,我们将matK、rbcL和trnL-intron获得的物种信息表利用Usearch进行标准化,标准化后数据相对丰度大于2(含)的物种会进入下游的数据分析;将各基因中分类等级门水平按标准化后数据量比例对三个基因的物种信息表进行相应比例的合并。
- 植物物种多样性分布格局
植物物种在所有样品中的整体分布进行总结后利用GraPhlan v1.1.3进行可视化。植物物种多样性在各样品中的分布情况利用Excel2016进行总结和可视化并利用Adobe Illustrator CC 2018进行美化,采用QGIS v3.6.3-Noosa绘制了植物物种多样性分布地图,以了解该地区的植物物种分布趋势。
- 植物物种多样性及其潜在驱动因素间的关联分析
本研究采用R v4.2.1对本地野生植物多样性与潜在驱动因子进行关联分析。气候因素与从野生植物物种数据集获得的α多样性指数之间进行了关联分析, 这些气候因子涵盖了温度、降水和其他相关因子,多样性指数包含了Shannon.e、Chao1、Berger Parker、Buzas Gibson、Dominance、Equitability、Jost、Richness和 Simpson等。本研究基于从Usearch alpha_div命令获得的α多样性指数,使用R v4.2.1对从土壤DNA获得的植物物种结果与地表植物物种调查结果进行了关联分析。使用Cytoscape v3.9.1对所有关联分析的结果进行可视化。
- 植物物种多样性与人类主要聚居地之间的UPGMA分析
使用R v4.2.1检验了人口密度和本地植物物种多样性之间的关联性并进行了UPGMA聚类分析。
引文格式:
Liu, Yanlei, Chao Xu,Wenpan Dong, Xun Chen, Wen Zhang, Yuzhe Sun, Guohong Wang, Yufei Wang, and Shiliang Zhou. 2023. "What determines plant species diversity along the Modern Silk Road in the east?" iMeta.e74. https://doi.org/10.1002/imt2.74
作者简介

刘艳磊(第一作者)
● 河北工程大学园林与生态工程学院讲师
● 目前研究方向为宏基因组学、分子生态学、进化生物学、植物基因组学、物种精准鉴定、物证溯源鉴定,相关学术成果已发表于iMeta、Molecular Phylogenetics and Evolution、Journal of Systematics and Evolution、BMC Plant Biology、Frontiers in Plant Science、Ecology and Evolution、BMC Genomics、Forensic Science International、Scientific Reports等期刊,参与国家自然科学基金多项,主持河北省自然科学基金青年基金一项

周世良(通讯作者)
● 中国科学院植物研究所研究员,博士生导师
● 研究方向为植物分类学、进化生物学、物种鉴定生物学、比较基因组学,已在Journal of Systematics and Evolution、Molecular Phylogenetics and Evolution、Molecular Ecology Resources、Proceedings of the Royal Society B: Biological Sciences、BMC Genetics、Journal of Integrative Plant Biology、American Journal of Botany、Proceedings of the National Academy of Sciences、iMeta等期刊发表学术论文百余篇,主持国家自然科学基金项目、中科院先导A项目子课题等多项,获得国家发明专利多项。