Contribution of CoA Ligases to Benzenoid Biosynthesis in Petunia Flowers
辅酶 A 连接酶在矮牵牛花卉苯环类物质生物合成中的作用

摘要
苯丙氨酸合成苯甲酸的过程中,需将侧链脱去两个碳原子,该过程可通过β- 氧化途径 或非氧化途径 完成。β- 氧化途径的起始步骤为生成肉桂酰辅酶 A,该反应大概率由 **4 - 香豆酸辅酶 A 连接酶(4CL)** 家族蛋白催化,这类酶可将多种反式肉桂酸衍生物转化为对应的辅酶 A 硫酯。
本研究利用功能基因组学手段,从矮牵牛花瓣特异性 cDNA 文库中筛选得到两个候选辅酶 A 连接酶基因。二者编码蛋白的氨基酸序列同源性仅为 25%,且均在矮牵牛花冠中高量表达。重组蛋白体外酶活鉴定结果显示:其中 Ph-4CL1 底物谱广泛,属于典型的 4 - 香豆酸辅酶 A 连接酶;另一个蛋白为肉桂酸辅酶 A 连接酶(Ph-CNL)。
RNA 干扰沉默实验表明,抑制Ph-4CL1 表达不会改变矮牵牛苯环类香气物质组成,而下调Ph-CNL 表达则会显著降低苯甲酸苯甲酯、苯乙基苯甲酸酯与苯甲酸甲酯的释放量。亚细胞定位实验显示,Ph-4CL1 定位于细胞质基质,Ph-CNL 定位于过氧化物酶体。
综上研究证实,酶蛋白的亚细胞定位模式决定其在苯环类物质合成通路中的功能分工;同时明确,过氧化物酶体内由 Ph-CNL 催化生成肉桂酰辅酶 A,是 β- 氧化途径中不可逆的关键限速步骤 。
引言
苯甲酸及其衍生物是植物体内各类化合物合成的重要结构单元,参与合成的物质范围十分广泛,既包括芳香类细胞分裂素、植物通用激素水杨酸等初生代谢物,也包含紫杉醇等具备化疗活性的物种特异性次生代谢产物。
大量含苄基(源自苯甲醇)或苯甲酰基(源自苯甲酸)结构的特化代谢物,在植物生命周期中也发挥着关键生态功能:既可作为防御物质抵御外界侵害,也能充当信号物质吸引传粉者与种子传播媒介。挥发性苯环类物质决定了众多植物的花香气味,也是植物挥发性代谢物中种类最丰富的大类之一。
研究普遍认为,苯甲酸是生物与非生物胁迫条件下水杨酸合成的前体物质;不过近年研究证实,在受病原菌侵染的拟南芥与本氏烟草中,还存在一条不依赖苯丙氨酸、直接以异分支酸为原料合成水杨酸的替代通路。
尽管植物苯环类物质分布广泛且生理功能重要,但植物体内苯甲酸的合成代谢通路目前仍未被完整阐明。 植物依托苯丙氨酸途径合成苯甲酸时,需要脱去侧链上两个碳原子,该过程主要存在三条代谢路径: 其一为β- 氧化途径 ,依次生成四类辅酶 A 硫酯中间产物,代谢流程为:反式肉桂酸→肉桂酰辅酶 A→3 - 羟基 - 3 - 苯基丙酰辅酶 A→3 - 氧代 - 3 - 苯基丙酰辅酶 A→苯甲酰辅酶 A→苯甲酸; 其二为非氧化裂解途径 ,先将水合态反式肉桂酸发生侧链裂解,生成中间产物苯甲醛,苯甲醛再进一步氧化生成苯甲酸; 第三条为非氧化途径的衍生通路,以肉桂酰辅酶 A 替代肉桂酸作为起始底物合成苯甲醛,最终生成苯甲酸。
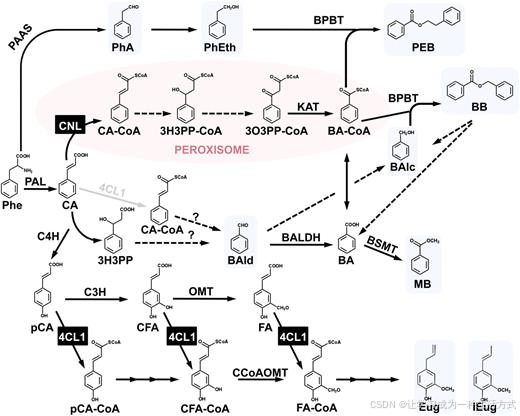
图 1 矮牵牛花瓣中苯甲酰辅酶 A 及苯环类 / 苯丙烷类挥发性物质推测生物合成通路
实心箭头代表已明确的生化反应,虚线箭头代表尚未阐明的潜在反应步骤;叠层箭头表示该过程包含多个酶促反应,灰色箭头代表推测反应步骤。 蓝色底色标注挥发性苯环类 / 苯丙烷类代谢物;参与合成苯甲酰辅酶 A、依赖辅酶 A 的 β- 氧化途径定位于过氧化物酶体,以粉色底色标注。
缩写释义 BA-CoA:苯甲酰辅酶 A;BAlc:苯甲醇;BAld:苯甲醛 BALDH:苯甲醛脱氢酶;BB:苯甲酸苯甲酯 BPBT:苯甲酰辅酶 A - 苯甲醇 / 2 - 苯乙醇苯甲酰转移酶 BSMT:苯甲酸 / 水杨酸羧基甲基转移酶 CA:肉桂酸;CA-CoA:肉桂酰辅酶 A;CFA:咖啡酸 CFA-CoA:咖啡酰辅酶 A;C3H:对香豆酸 - 3 - 羟化酶 C4H:肉桂酸 - 4 - 羟化酶;CCoAOMT:咖啡酰辅酶 A 3-O - 甲基转移酶 4CL1:4 - 香豆酸辅酶 A 连接酶;Eug:丁子香酚 FA:阿魏酸;FA-CoA:阿魏酰辅酶 A;IEug:异丁子香酚 3H3PP:3 - 羟基 - 3 - 苯基丙酸;3H3PP-CoA:3 - 羟基 - 3 - 苯基丙酰辅酶 A KAT:3 - 酮脂酰辅酶 A 硫解酶;MB:苯甲酸甲酯 3O3PP-CoA:3 - 氧代 - 3 - 苯基丙酰辅酶 A;OMT:氧 - 甲基转移酶 PAAS:苯乙醛合酶;pCA:对香豆酸;pCA-CoA:对香豆酰辅酶 A PEB:苯乙基苯甲酸酯;PhA:苯乙醛;PhEth:2 - 苯乙醇
所有苯甲酸合成通路均起始于 ** 苯丙氨酸解氨酶(PAL)** 催化 L - 苯丙氨酸生成反式肉桂酸,后续通路开始分支。β- 氧化途径的起始反应为合成肉桂酰辅酶 A,该反应推测由 4 - 香豆酸辅酶 A 连接酶家族(4CL,EC 6.2.1.12)成员催化。 4CL 酶普遍底物谱较广,可催化多种肉桂酸羟基 / 甲氧基衍生物生成对应的辅酶 A 硫酯。但目前已鉴定的绝大多数植物 4CL 难以高效利用反式肉桂酸作为底物,仅有树莓 4CL2 对反式肉桂酸亲和力极强,酶活效率可达香豆酸底物的 153%,迄今尚未发现专一识别肉桂酸的肉桂酸辅酶 A 连接酶(CNL)。
本研究选用矮牵牛花瓣作为研究材料,探究植物体内苯环类物质合成机制。矮牵牛花瓣可大量释放苯甲酸甲酯(苯甲酸甲基化产物),以及多种含苯甲酰基、苄基结构的挥发性香气物质。 课题组前期已鉴定并解析了催化这类香气物质合成的关键酶,包括苯甲酸甲酯、苯甲酸苯甲酯、苯乙基苯甲酸酯合成相关酶;同时结合代谢流量分析与活体稳定同位素标记技术证实,β- 氧化途径与非氧化途径共同参与矮牵牛苯环类香气物质合成 。本研究进一步分离鉴定 β- 氧化途径第一步关键酶肉桂酸辅酶 A 连接酶,并证实该步反应定位于过氧化物酶体中完成。
结果
催化肉桂酰辅酶 A 合成的候选辅酶 A 连接酶基因筛选
为挖掘参与肉桂酰辅酶 A 合成的功能基因,本研究以拟南芥 4CL1 序列为参照,设置阈值 e<sup>-10</sup>,在矮牵牛花瓣特异性表达序列标签数据库及茄科基因组数据库中开展 tBLASTn 同源序列检索。
从花瓣特异数据库中筛选得到两类同源序列:第一类 4 条 EST 序列对应同一基因,编码蛋白与植物已知 4CL 同源性高;第二类 4 条 EST 序列对应另一基因,其编码蛋白与 4CL 同源性较低,但隶属于酰基激活酶超家族。该家族蛋白催化机制保守,可先形成腺苷酸化底物中间体,再完成辅酶 A 酯化反应。 结合后续功能验证结果,将前者命名为Ph-4CL1 ,后者命名为Ph-CNL (矮牵牛肉桂酸辅酶 A 连接酶基因)。 从茄科基因组数据库中还检索到 3 条与植物 4CL 高度同源的 EST 序列。
苯环类香气物质主要在矮牵牛花冠中合成释放,因此香气合成相关基因大多在花冠中优势表达。研究采用基因特异性引物,通过实时荧光定量 PCR,对 5 个候选基因在开放 2 天的矮牵牛花冠组织中进行表达模式检测。 结果显示,其中 3 个基因在花冠裂片内表达量极低甚至不表达,表达规律与花香物质合成无关,不再进行后续功能研究; 而Ph-4CL1 与Ph-CNL 在花冠中特异性高表达,进入深入分析阶段。
矮牵牛花冠中苯丙烷类、苯环类物质内源积累量与挥发性释放量存在显著昼夜节律变化,与之相符,Ph-4CL1 与Ph-CNL 在花冠裂片内的表达量同样呈现昼夜节律,表达峰值集中在每日傍晚 15:00---19:00; 在花朵不同发育阶段,两个基因的表达趋势高度一致,表达水平与苯环类香气物质合成积累量呈正相关。
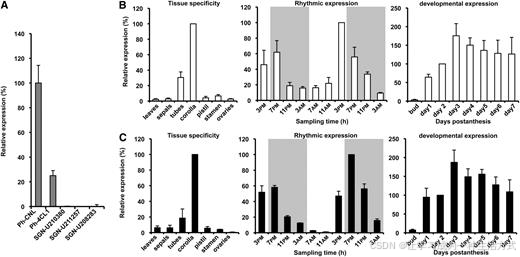
图 2 矮牵牛中候选 4CL 基因与 CNL、4CL1 基因的表达模式
所有基因转录水平均通过实时荧光定量 PCR 检测,以延伸因子 1-α 为内参基因进行相对定量;数据为平均值 ± 标准误差,设置 3 组生物学重复。
(A) 选取开花后第 2 天下午 3 点的花冠组织,检测 SGN-U210380、SGN-U211257、SGN-U208283、CNL 、4CL1 的表达量,以表达量最高的CNL 为参照进行相对表达量统计。
(B)(C) 左侧图:分别以花冠组织中的转录本丰度为参照,展示CNL 与4CL1 在不同组织中的特异性表达特征; 中间图:统计开花后 1~3 天内,自然昼夜周期下花冠中CNL 与4CL1 的 mRNA 节律表达变化,CNL 以开花后第 2 天 15:00 表达量为参照,4CL1 以同日 19:00 表达量为参照;灰色区域代表黑暗期,白色区域代表光照期; 最右侧图:统计从成熟花蕾至开花后第 7 天,花冠内两个基因在花朵不同发育阶段的表达变化,统一以开花后第 2 天的表达水平作为参照基准。
为进一步预判并区分Ph-4CL1 与Ph-CNL 的生物学功能,本研究以已分类的拟南芥酰基激活酶(AAE)超家族成员为序列比对框架,结合同源检索结果与已完成生化功能鉴定的同源酶蛋白序列构建系统发育树。
邻接进化树结果显示,Ph-4CL1 与 Ph-CNL 分属完全不同的进化亚家族 (图 3)。 Ph-4CL1 归属于第 IV 进化分支,该分支以 4 - 香豆酸辅酶 A 连接酶为主;其氨基酸序列与烟草 4CL1、4CL2 同源性约 90%,与 3 种已鉴定的拟南芥 4CL 酶同源性约 70%。 而 Ph-CNL 归属于第 VI 进化分支,与古代稀的苯甲酰辅酶 A 连接酶 / 苯甲酰氧基芥子油苷合成酶亲缘关系最近(序列一致性 61%、相似性 74%),该植物花瓣可大量释放苯环类香气物质;同时与拟南芥同源蛋白 At1g65880 相似度较高(一致性 60%、相似性 73%),已有研究证实该拟南芥蛋白具备苯甲酸辅酶 A 连接酶活性。
综上,系统发育分析表明Ph-CNL 与Ph-4CL1 功能存在明显分化;结合其与古代稀 BZL 蛋白、拟南芥 BZO1 蛋白的进化亲缘关系可推测,Ph-CNL 极有可能参与苯环类物质生物合成的 β- 氧化代谢通路 。
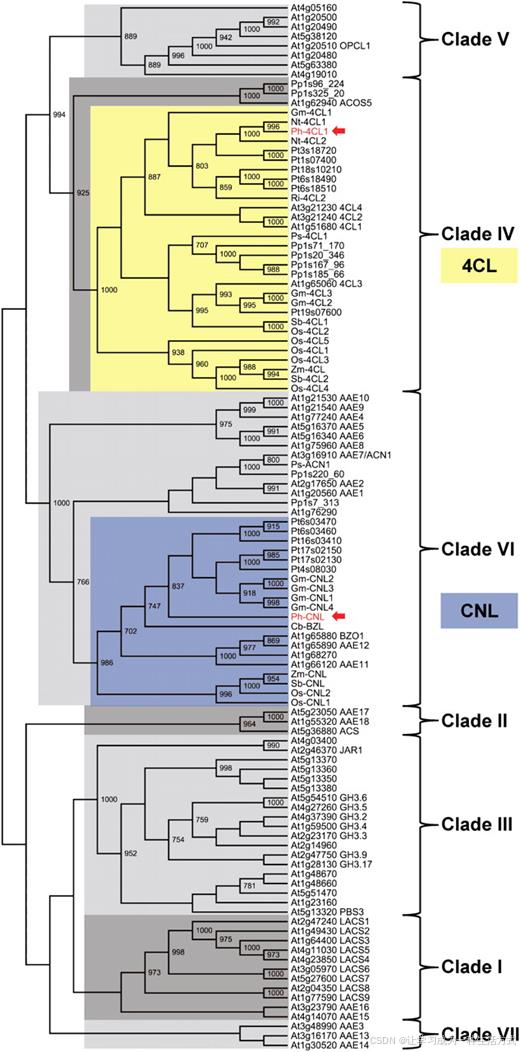
图 3 Ph-4CL1 与 Ph-CNL 分属 Shockey 和 Browse(2011)划分的酰基激活酶超家族不同进化亚家族
通过非穷尽式蛋白序列比对检索,筛选获得更多 Ph-4CL1 与 Ph-CNL 同源蛋白序列;利用 ClustalO 1.0.3 软件完成多序列比对。 基于比对结果,借助 PHYLIP 3.69 软件构建邻接系统发育分支树;分支节点处标注自举检验值(重复抽样 1000 次),仅展示支持度≥70% 的数值。序列登录号详见附表 1。
物种缩写 At:拟南芥;Cb:古代稀;Gm:大豆;Nt:烟草;Os:水稻; Ph:矮牵牛;Pp:小立碗藓;Ps:北美云杉;Pt:毛果杨; Ri:树莓;Sb:高粱;Zm:玉米
Ph-4CL1 与 Ph-CNL 蛋白功能鉴定
为明确两种候选辅酶 A 连接酶的生化功能,将两个基因的编码区序列导入大肠杆菌,诱导表达带有六组氨酸标签 的融合重组蛋白。 经凝胶过滤色谱测定重组蛋白分子量,并与理论分子量比对证实:两种有活性的酶蛋白均以单体形式 存在,其中 Ph-4CL1 分子量 59 kDa,Ph-CNL 分子量 63 kDa。
对亲和层析纯化后的重组蛋白开展底物活性检测,底物涵盖肉桂酸及其多种羟基、甲氧基衍生物,测定其催化生成对应辅酶 A 硫酯的能力。 结果显示:两种酶均可催化肉桂酸、对香豆酸、咖啡酸生成辅酶 A 酯;但仅 Ph-4CL1 能够利用阿魏酸与苯甲酸作为催化底物(表 1)。 酶活最适 pH 测定结果表明:以肉桂酸为底物时,Ph-4CL1 最适 pH 为 8.5,Ph-CNL 最适 pH 为 8.0。
|----------------|--------------------------|-----------------------------|-----------------------------------------------|-----------------------------------------|-------------------------------------------------------------------------------|
| Enzyme | Substrate (Acid) | K m****(µM)**** | V max****(pkat mg**** −1****)**** | k cat****(s**** −1****)**** | k cat****/**** K m****(mM**** −1s −1****)**** |
| Ph-4CL1 | t-Cinnamic | 149.4 ± 13.7 | 14680 ± 409 | 0.847 ± 0.02 | 6.43 ± 0.86 |
| | 4-Coumaric | 16.4 ± 3.0 | 25797 ± 1994 | 1.488 ± 0.12 | 84.32 ± 9.00 |
| | Caffeic | 47.8 ± 5.4 | 63200 ± 2020 | 3.645 ± 0.12 | 70.14 ± 5.95 |
| | Ferulic | 15.2 ± 2.7 | 41377 ± 879 | 2.39 ± 0.05 | 145.37 ± 27.21 |
| | Benzoic | 9008 ± 1795 | 95.9 ± 8.5 | 0.0007 ± 0.0001 | 0.0008 ± 0.0001 |
| | CoA | 9.9 ± 3.1 | 8451 ± 4017 | 0.487 ± 0.23 | 49.46 ± 6.51 |
| Ph-CNL | t -Cinnamic | 285.7 ± 63.9 | 7401.7 ± 117.2 | 0.472 ± 0.01 | 1.82 ± 0.36 |
| | 4-Coumaric | 550.4 ± 127.5 | 3890.7 ± 460.1 | 0.248 ± 0.03 | 0.512 ± 0.14 |
| | Caffeic | 1042.4 ± 95.9 | 3742.7 ± 3.6 | 0.238 ± 0 | 0.232 ± 0.02 |
| | Ferulic | n.d. | n.d. | n.d. | n.d. |
| | Benzoic | n.d. | n.d. | n.d. | n.d. |
| | CoA | 775.2 ± 34.3 | 7179.7 ± 126.8 | 0.458 ± 0.01 | 0.607 ± 0.06 |
测定 Ph-4CL1 的酶促动力学参数发现:该酶对对香豆酸 与阿魏酸 的表观米氏常数\(K_m\)数值相近,分别为 16.4 μM、15.2 μM,约为咖啡酸(47.8 μM)的三分之一;其对肉桂酸的表观\(K_m\)为 149.4 μM。该酶也可催化苯甲酸生成对应的硫酯,但对苯甲酸亲和力极低,表观\(K_m\)高达 9 mM。
催化效率(\(k_{cat}/K_m\))分析显示,Ph-4CL1 对阿魏酸催化效率最高,其次为对香豆酸与咖啡酸;其对肉桂酸的催化效率不足阿魏酸的 1/20。Ph-4CL1 可高效催化对香豆酸、咖啡酸、阿魏酸生成对应辅酶 A 酯,证实该蛋白是典型的 4 - 香豆酸辅酶 A 连接酶。
对 Ph-CNL 进行动力学参数测定结果表明:该酶对底物亲和力表现为肉桂酸>对香豆酸>咖啡酸,三者表观\(K_m\)依次为 285 μM、550 μM、1042 μM;以肉桂酸为底物时的催化效率,分别是对香豆酸的 3.6 倍、咖啡酸的 8 倍,说明反式肉桂酸是其最适底物 ,因此将该酶正式命名为肉桂酸辅酶 A 连接酶(CNL)。
经飞行时间液相色谱 - 质谱联用(LC/TOF-MS)鉴定,Ph-CNL 与 Ph-4CL1 催化肉桂酸生成的产物,在分子量与色谱保留行为上,均与烟草 4CL 体外合成的肉桂酰辅酶 A 标准品完全一致。
Ph-CNL 催化肉桂酸的活性严格依赖钾离子:体系中氯化钾浓度为 10.5 mM 时,酶活可达最大活性的半数;若蛋白纯化与酶活反应体系中完全去除钾离子,该酶催化肉桂酸的活性会彻底丧失,且仅能通过补加钾离子恢复活性,钠离子无恢复作用,仅可恢复约 24% 酶活。
已有研究报道,绿弯菌 3 - 羟基丙酸辅酶 A 合成酶、醋杆菌乙酰辅酶 A 合成酶、拟青霉丁酰辅酶 A 合成酶均为镁离子 + 钾离子 双重依赖型酶,与 Ph-CNL 特性一致。缺失钾离子也会显著抑制 Ph-CNL 催化对香豆酸的活性(降幅 73%),对咖啡酸催化活性影响相对较小(降幅 14%)。
两种酶底物催化能力差异极大:Ph-4CL1 催化对香豆酸、咖啡酸的效率,分别是 Ph-CNL 的 165 倍、305 倍;即便肉桂酸是 Ph-CNL 最适底物,其催化效率也仅为 Ph-4CL1 的 1/3.5。此外,Ph-CNL 对辅酶 A 的亲和力远低于 Ph-4CL1,二者对应的辅酶 A 表观\(K_m\)分别为 775 μM、9.9 μM。
RNA 干扰沉默 4CL1 与 CNL 对苯环类物质合成的影响
仅凭基因表达模式与体外酶活底物特性,仍无法明确两种酶在矮牵牛花瓣挥发性苯环类物质合成中的体内功能。因此本研究以米切尔矮牵牛为材料,利用RNA 干扰技术 分别下调两个基因表达,解析二者生理功能。
分别截取4CL1 、CNL 编码区 5' 端 431 bp、473 bp 的基因特异性片段构建干扰载体,借助花瓣特异性 LIS 启动子驱动干扰片段表达。最终获得 25 株4CL1 干扰株系、16 株CNL 干扰株系。
筛选目标基因表达下调幅度超 60% 的阳性株系,系统测定花瓣释放的挥发性香气物质、体内中间代谢物与终产物含量。实验证实:沉默CNL 不会影响4CL1 表达,反之亦然,二者不存在表达互扰现象。
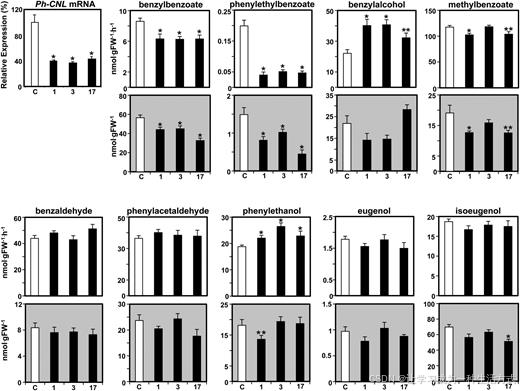
图 4 矮牵牛花冠中 RNA 干扰沉默 CNL 对基因表达、苯环类 / 苯丙烷类物质释放量及内源积累量的影响
于开花后第 2 天下午 3 点采集花冠组织,通过实时荧光定量 PCR 检测对照组(白色柱形)与 3 个独立CNL 干扰株系(1、3、17 号,黑色柱形)的CNL 转录水平;以对照组花瓣CNL 表达量为 100%,统计转基因株系相对表达比例,数据为平均值 ± 标准误差,设 3 组生物学重复。
白色背景图为挥发性物质释放速率,灰色背景图为对应物质的内源积累含量。 选取开花后第 2 天离体花朵,自晚间 8 时开始持续收集 12 小时花香组分,假定 12 小时内释放速率恒定,换算为每小时释放量;内源代谢物样品统一在晚间 8 时取样。数据为平均值 ± 标准误差,生物学重复数≥10;采用独立样本 t 检验分析差异,*P<0.05、**P<0.01;FW 代表鲜重。
干扰下调CNL 表达后,依赖苯甲酰辅酶 A 合成的苯甲酸苯甲酯、苯乙基苯甲酸酯 平均释放量分别下降约 27%、77%; 而合成这两种酯类的另一类酰基受体底物 ------ 苯甲醇、2 - 苯乙醇的释放量分别上调 72%、27%。
CNL 干扰植株的苯甲酸甲酯释放量整体显著下降 12%,仅 3 号株系与对照组无明显差异;苯甲醛、苯乙醛、丁子香酚、异丁子香酚的释放量均未发生改变。
整体来看,转基因株系与野生型的挥发性物质释放量,大多与其组织内源积累量呈正相关;但苯甲醇与 2 - 苯乙醇例外,二者释放量升高,内源含量并未同步上升。
对关键酶活测定结果显示:催化合成苯甲酸苯甲酯 / 苯乙基苯甲酸酯的苯甲酰转移酶、催化合成苯甲酸甲酯的羧酸甲基转移酶,以及其他花香合成相关酶、4CL 酶活性,在CNL 干扰花冠中均与对照组无显著差异。
沉默CNL 不会影响游离羟基肉桂酸的内源含量,但会使 β- 氧化途径终产物苯甲酰辅酶 A 平均含量下降 38%,其余各类羟基肉桂酸辅酶 A 酯的含量均无明显变化。
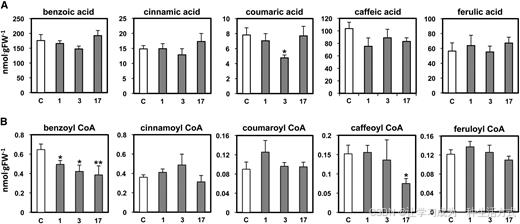
图 5 矮牵牛花冠中沉默 CNL 对游离羟基肉桂酸及其辅酶 A 酯内源含量的影响
于开花后第 2 天晚 8 点采集对照组(白色柱)与CNL 干扰株系(灰色柱)花冠组织,分别提取游离有机酸(A)与羟基肉桂酰辅酶 A 酯类物质(B)。 数据以平均值 ± 标准误差表示,生物学重复≥6 组;采用 t 检验进行差异分析,**P<0.04,*P<0.01。C 为对照组,FW 表示鲜重。
将4CL1 基因表达量通过 RNA 干扰下调 60% 后,转基因花瓣粗酶液中 Ph-4CL1 酶活性同步下降 61%,对照组酶活为 38.26±7.35 皮卡托 / 毫克蛋白,转基因株系仅为 15.05±4.15 皮卡托 / 毫克蛋白。 酶活测定选用阿魏酸作为底物,以此区分两种酶活性,原因是 Ph-CNL 无法催化利用阿魏酸。下调4CL1 转录水平后,植株各类苯环类挥发性物质的释放量均未出现明显变化。
Ph-4CL1 与 Ph-CNL 蛋白的亚细胞定位
利用蛋白定位预测软件 PSORT 进行分析,结果显示:Ph-CNL 蛋白 C 端含有典型的过氧化物酶体定位信号 PTS1(AKL) ,预测定位于过氧化物酶体;而 Ph-4CL1 无此类定位信号,预测定位于细胞质基质。
为明确其实际亚细胞定位,将含完整 PTS1 定位信号的 CNL 编码序列 融合至绿色荧光蛋白(GFP)编码区的 3' 端,构建融合表达载体;随后转入拟南芥原生质体,借助激光共聚焦显微镜观察瞬时表达的荧光分布特征(图 6)。
荧光观察结果显示,GFP-Ph-CNL 融合蛋白呈现点状荧光分布,符合过氧化物酶体蛋白的定位特征;通过远红光滤光片检测 637 nm 处叶绿体自发荧光并以蓝色标注叠加成像,证实该融合蛋白并未定位于叶绿体 。
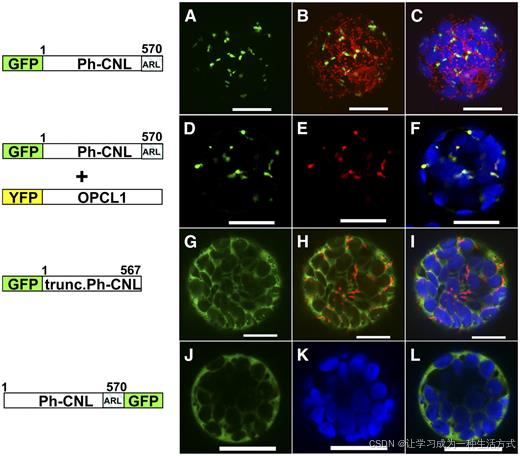
图 6 Ph-CNL 蛋白的亚细胞定位
左侧为绿色荧光蛋白(GFP)融合载体构建示意图,右侧为激光共聚焦显微镜观察得到的拟南芥原生质体瞬时表达结果。 绿色代表 GFP 荧光信号,蓝色为叶绿素自发荧光;除 E 图外,其余实验组均采用线粒体红色荧光探针标记线粒体(红色荧光),以此区分线粒体与过氧化物酶体;E 图中以定位于过氧化物酶体的标记蛋白 OPCL1 的黄色荧光蛋白(YFP)荧光作为红色参照。 叠加图为 GFP、线粒体、叶绿体与过氧化物酶体荧光信号的合并成像;载体示意图中的数字代表氨基酸位点,ARL 为 Ph-CNL 蛋白所含的过氧化物酶体定位信号肽。每组载体仅展示两次独立转化实验中的其中一组结果,比例尺 = 50 微米。
为区分过氧化物酶体与线粒体、排除荧光信号定位于线粒体的可能性,对表达 GFP-Ph-CNL 融合蛋白的原生质体进行线粒体红色荧光探针染色。结果显示,GFP 绿色荧光与线粒体红色荧光无重叠,证明Ph-CNL 不定位在线粒体 。
同时,Ph-CNL 荧光信号与已证实定位于过氧化物酶体的拟南芥酰基激活酶 OPCL1 荧光信号完全共定位,进一步确定 Ph-CNL 定位于过氧化物酶体。
为验证 Ph-CNL 蛋白 C 端的PTS1 型定位基序 ARL 是决定其亚细胞定位的核心元件,构建去除 C 端 ARL 三肽的截短型 Ph-CNL 并与 GFP 融合表达。结果发现,缺失 ARL 序列后,蛋白定位由过氧化物酶体转为细胞质基质; 若将 GFP 连接在 Ph-CNL 蛋白 C 端,会直接遮挡 PTS1 定位信号,同样致使融合蛋白丧失过氧化物酶体定位能力,最终定位于细胞质基质。
与之相反,无论将 GFP 连接在 Ph-4CL1 蛋白的 N 端还是 C 端,该融合蛋白均稳定定位于细胞质基质,与前期预测结果一致。
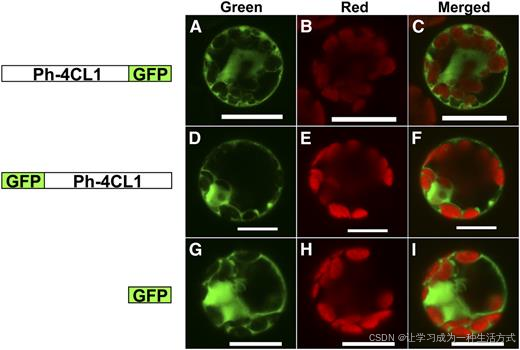
图 7 矮牵牛 4CL1 蛋白的亚细胞定位
左侧为 GFP 融合表达载体结构示意图,右侧为原生质体瞬时表达的显微观察结果。绿色通道显示 GFP 荧光信号,红色通道显示叶绿体自发荧光,叠加图为 GFP 与叶绿体荧光的合并图像。 以 p326-sGFP 空载载体作为细胞质定位阳性对照,比例尺为 50 微米。
为从生化层面进一步证实 Ph-CNL 定位于过氧化物酶体,本研究提取矮牵牛花瓣过氧化物酶体组分并测定辅酶 A 连接酶活性。结果显示,与体外重组 Ph-CNL 酶活特征一致,花瓣过氧化物酶体提取物无法催化阿魏酸发生反应(阿魏酸是 4CL 特征底物); 且其催化肉桂酸的酶活是对香豆酸的 2.4 倍,该比值与重组 Ph-CNL 两种底物最大反应速率比值(1.9 倍)高度吻合。 相较于野生型,CNL 干扰株系花瓣过氧化物酶体内 CNL 酶活性下降 67%,与该基因转录水平下调 60% 的结果相符。
鉴于 Ph-CNL 定位于过氧化物酶体,本研究还探究其是否参与脂肪酸 β- 氧化过程。利用重组 Ph-CNL 分别检测短链、中链、长链脂肪酸底物活性,结果表明该酶几乎无法催化各类脂肪酸生成对应的辅酶 A 酯。
讨论
矮牵牛 4CL1 与 CNL 酶的异同特征
植物体内苯甲酸的合成通路至今仍未被完全阐明,多项研究证实过氧化物酶体 β- 氧化途径 参与植物苯甲酸合成。例如,拟南芥chy1 突变体的过氧化物酶体 β- 氧化过程受阻,种子中苯甲酰化芥子油苷含量显著降低;矮牵牛花瓣中过氧化物酶体酮脂酰硫解酶基因表达下调后,依赖苯甲酰辅酶 A 合成的挥发性香气物质含量也随之下降。
以苯丙氨酸为前体合成苯甲酸的 β- 氧化途径中,生成肉桂酰辅酶 A 是首个关键限速步骤 ,该反应必须依靠辅酶 A 连接酶催化。目前已在拟南芥、烟草、大豆、欧芹、杨树、树莓等多种植物中鉴定出典型 4CL 酶,这类酶可高效催化各类羟基、甲氧基取代型肉桂酸生成辅酶 A 硫酯,但绝大多数 4CL 难以利用肉桂酸作为底物,仅树莓 4CL2 对肉桂酸具备偏好性。
本研究在矮牵牛花瓣中筛选得到 5 个候选辅酶 A 连接酶基因,其中Ph-4CL1 与Ph-CNL 的表达模式与花香类苯环物质合成规律高度契合。二者均能催化肉桂酸生成肉桂酰辅酶 A,但体外催化效率存在差异; Ph-4CL1 可高效利用阿魏酸、对香豆酸、咖啡酸,对肉桂酸催化活性偏弱;而 Ph-CNL专一偏好肉桂酸,完全无法利用阿魏酸 ,这一底物偏好性使其与经典 4CL 酶明确区分。
两种酶底物谱存在部分重叠、酶学性质差异显著,且氨基酸序列同源性仅 25%;系统发育分析显示二者分属酰基激活酶超家族下两个完全独立的进化分支,属于功能趋同进化 产物。 Ph-4CL1 隶属于功能研究透彻的 4CL 进化分支,主要负责各类苯丙烷类物质合成前体的活化;Ph-CNL 则归属于功能大多未知的植物特有辅酶 A 连接酶分支。 Ph-CNL 的同源蛋白为古代稀 BZL 蛋白与拟南芥 BZO1 蛋白,已有研究证实拟南芥 BZO1 可催化苯甲酸生成苯甲酰辅酶 A,结合本研究发现的钾离子依赖特性,后续有必要重新验证 At-BZO1 对肉桂酸的催化活性。
4CL 与 CNL 两类蛋白并非矮牵牛独有,在单子叶、双子叶植物中广泛存在;但苔藓、云杉等低等陆生植物中的同源蛋白更贴近乙酰辅酶 A 连接酶,推测CNL 蛋白为开花植物特有 ,非开花植物若依靠 β- 氧化途径合成苯甲酸,则不依赖 CNL 酶发挥作用。
过氧化物酶体定位的 CNL 介导苯甲酸 β- 氧化合成通路首个限速步骤
两种酶最核心的差异在于亚细胞定位 :Ph-4CL1 定位于细胞质基质,Ph-CNL 定位于过氧化物酶体。 Ph-CNL 对辅酶 A 的亲和力远低于 Ph-4CL1,暗示过氧化物酶体内游离辅酶 A 含量充足,与该细胞器是脂肪酸 β- 氧化主要场所、乙酰辅酶 A 富集的特征相符。 同时 Ph-CNL 对肉桂酸整体底物亲和力偏低,而植物体内尚未发现定位于过氧化物酶体的苯丙氨酸解氨酶,由此推测植物体内存在专门将肉桂酸转运进入过氧化物酶体的转运机制,以此保障 Ph-CNL 高效催化反应进行。
二者表达模式相近,均可活化肉桂酸,且 Ph-4CL1 体外催化效率更高,但体内功能截然不同:下调Ph-CNL 表达会显著降低苯甲酸苯甲酯、苯乙基苯甲酸酯的释放量,下调Ph-4CL1 则对花香物质合成无明显影响。 上述两种香气物质均以苯甲酰辅酶 A 为直接前体,而CNL 干扰株系花瓣内苯甲酰辅酶 A 含量平均下降 38%,直接印证体内功能差异。 干扰株系中肉桂酸与肉桂酰辅酶 A 总含量无明显变化,原因在于细胞质中 Ph-4CL1 同样可催化生成肉桂酰辅酶 A,常规检测无法区分细胞质与过氧化物酶体内的组分含量。
矮牵牛花香代谢流量分析表明,植物体内苯甲酰辅酶 A 主要由 β- 氧化途径合成,仅有少量来源于非氧化途径生成的苯甲酸;同位素示踪实验也证实了这条次要代谢支路的存在。 沉默CNL 后,植物代谢流可转向非氧化途径合成苯甲酸,弥补部分苯甲酰辅酶 A 供给,这也解释了为何 CNL 酶活下降 67%,而体内苯甲酰辅酶 A 含量仅下降 38%。
已有代谢调控研究表明,苯环类物质合成通路中,仅参与苯甲酸苯甲酯合成与降解的关键酶能够调控该物质代谢流量与释放量。β- 氧化途径中催化 3 - 氧代 - 3 - 苯基丙酰辅酶 A 生成苯甲酰辅酶 A 的硫解酶基因沉默后,同样会显著降低矮牵牛花瓣中两类苯环酯类香气物质含量。 结合 Ph-CNL 无法活化苯甲酸的酶活特征可确定:植物体内 Ph-CNL 仅在过氧化物酶体内催化生成肉桂酰辅酶 A,专一参与 β- 氧化途径合成苯甲酰辅酶 A 。
前人研究发现拟南芥BZO1 突变体种子中苯甲酰化芥子油苷合成受阻,体外酶活实验证明 At-BZO1 具备苯甲酸辅酶 A 连接酶活性,据此推测其在植物体内负责为 BAHD 酰基转移酶提供反应底物。 但该研究并未验证其对肉桂酸的催化能力,也未检测突变体种子内苯甲酸积累量;同时 At-BZO1 同样携带过氧化物酶体定位信号,而芥子油苷主要在细胞质中合成。 结合本研究结果可合理推测:拟南芥 At-BZO1 真实生理功能大概率是催化肉桂酸生成肉桂酰辅酶 A,参与 β- 氧化途径合成苯甲酸,进而为芥子油苷苯甲酰化修饰提供原料,该推论仍需体外酶活验证与突变体代谢组分析加以证实。
沉默CNL 同样会小幅下调苯甲酸甲酯释放量,但影响程度远弱于两类苯环酯类物质,降幅仅约 12%。该现象由两大因素共同导致:一是花瓣内苯甲酸苯甲酯可作为苯丙氨酸合成苯甲酸的中间产物,其含量下降间接影响苯甲酸积累;二是代谢流向非氧化途径分流,补充了苯甲酸的合成来源。
苯甲酰辅酶 A 在过氧化物酶体内合成,而以其为直接前体的苯甲酸苯甲酯、苯乙基苯甲酸酯均在细胞质中完成合成。目前在动物与酵母中已发现可转运酰基辅酶 A 酯进出过氧化物酶体的 ABC 转运蛋白家族,但植物体内尚未发现能够将酰基辅酶 A 酯运出过氧化物酶体的转运机制 。
综上,尽管 Ph-4CL1 与 Ph-CNL 对肉桂酸催化效率相近、表达模式一致,但仅定位于过氧化物酶体的 Ph-CNL 负责将肉桂酸导入 β- 氧化代谢通路,主导体内苯甲酰辅酶 A 及其衍生物的生物合成 。而过氧化物酶体内合成的苯甲酰辅酶 A 如何转运至细胞质,进而参与矮牵牛挥发性苯环香气物质合成,仍是后续亟待探究的关键问题。
材料与方法
植物材料
以米切尔矮牵牛为遗传背景,构建获得4CL1 与CNL 基因 RNA 干扰转基因株系。野生型对照植株与转基因植株分别栽培于常规温室环境与组培体系中,统一生长月龄与生理状态,用于后续各项实验。
系统发育分析
利用 ClustalO 软件完成氨基酸多序列比对;借助 PHYLIP 3.69 软件构建无根邻接进化树,设置自举检验重复次数为 1000 次,最终使用 FigTree 1.3.1 软件可视化进化树,所有程序均采用默认参数运行。
重组蛋白表达与纯化
扩增矮牵牛4CL1 完整编码区,上下游引物序列分别为 5′-AACCCATATGCCGATGGAGACTGAAAC-3′、5′-GGCATATGTTAATTTGGAACACCAGCTGC-3′,将其插入带有可切除 N 端六组氨酸标签的原核表达载体 pET-28a 的Nde Ⅰ 酶切位点处。
扩增Ph-CNL 编码区,上下游引物依次为 5′-ATCATATGGACGAGTTACCAAAATGTGGAGC-3′、5′-ATGCGGCCGCCAGACGAGCTGGCAAATCAAG-3′,精准插入 pET-32a 载体Nde Ⅰ-Not Ⅰ 位点,保证读码框完整且位于六组氨酸标签上游。测序验证 PCR 扩增序列无突变错误。
将重组质粒及空载对照质粒分别转入大肠杆菌 Rosetta 感受态细胞,pET-28a 载体菌株用含 50 μg/mL 卡那霉素、pET-32a 载体菌株用含 100 μg/mL 氨苄青霉素的 LB 液体培养基,37℃恒温培养。待菌体培养液\(A_{600}\)值达到 0.5 时,加入异丙基 -β-D - 硫代半乳糖苷至终浓度 0.4 mM 诱导蛋白表达;18℃、200 rpm 摇床振荡培养 16 h 后收集菌体。
重悬缓冲液配制:Ph-4CL1 菌体重悬液含 50 mM 磷酸钾(pH 8.0)、500 mM 氯化钠、10% 甘油、1 mM EDTA、7 mM β- 巯基乙醇;Ph-CNL 菌体重悬液含 50 mM 磷酸钾(pH 8.0)、150 mM 氯化钾、20% 甘油、1 mM 苯甲基磺酰氟。
向菌液中加入溶菌酶与脱氧核糖核酸酶 Ⅰ 冰上裂解 30 min,再进行超声破碎;4℃、20000 g 离心 30 min 获取上清粗酶液。采用 AKTA 快速蛋白液相色谱系统,借助镍离子亲和层析柱纯化目的蛋白,洗脱流程先用梯度咪唑溶液除杂,最后用 500 mM 咪唑洗脱融合蛋白。
收集高浓度蛋白洗脱液,经脱盐柱置换为含 20 mM 磷酸钾(pH 8.0)、20% 甘油的保存缓冲液;蛋白分装后液氮速冻,置于 - 80℃冰箱低温保存。经考马斯亮蓝染色 SDS - 聚丙烯酰胺凝胶电泳灰度分析,Ph-4CL1 蛋白纯度约 60%,Ph-CNL 纯度约 85%,计算催化常数时已校正纯度差异;蛋白浓度采用布拉德福德法测定。
酶活测定
采用放射性同位素法与紫外分光光度法同步测定辅酶 A 连接酶活性,两种检测方式结果一致。
放射性同位素法 :总反应体系 100 μL,Ph-CNL 反应体系含 2.7 μg 纯化蛋白、放射性标记肉桂酸,搭配 50 mM 复合缓冲液、ATP、氯化镁、氯化钾与辅酶 A;加入辅酶 A 启动反应,室温反应 20 min 后用三氯乙酸终止反应,乙酸乙酯反复萃取去除游离底物,取水相液进行液体闪烁计数,测定放射性掺入量。空白对照设置高温灭活酶液组与无酶体系组。Ph-4CL1 反应体系依据最适反应条件调整缓冲液 pH、离子浓度与底物用量。放射性标记肉桂酸由放射性苯丙氨酸经苯丙氨酸解氨酶体外催化合成。
紫外分光光度法 :总体系 500 μL,反应基础条件与放射性法一致,替换为非放射性底物,加入不同羟基肉桂酸底物启动反应。依据各类辅酶 A 酯特征吸收峰波长实时监测吸光度变化,结合对应摩尔消光系数计算酶促反应生成产物量。
苯甲酸底物酶活检测同样采用放射性法,调整反应缓冲液 pH 与离子配比,测定 Ph-4CL1 催化活性。
长短链脂肪酸底物活性采用偶联酶体系测定,通过检测反应体系中 AMP 生成量,结合还原型辅酶 Ⅰ 氧化速率表征酶活;脂溶性脂肪酸底物先用含曲拉通 X-100 的二甲基亚砜助溶。
参照已发表实验方法,测定野生型与转基因植株开花后第 2 天花瓣粗酶液中苯丙氨酸解氨酶、苯甲醛脱氢酶、异丁子香酚合酶、丁子香酚合酶、羧酸甲基转移酶、苯甲酰转移酶、苯乙醛合酶等花香合成通路关键酶活性。
液相色谱 - 质谱联用产物鉴定
利用高效液相色谱串联飞行时间质谱仪鉴定酶促反应产物,以烟草 4CL 体外催化生成的肉桂酰辅酶 A 作为阳性对照。色谱条件:采用 C18 色谱柱,柱温 40℃,流动相为乙酸铵水溶液 - 乙酸铵甲醇溶液体系,设置线性洗脱梯度;质谱采用电喷雾负离子模式,优化离子源温度、鞘气流速、毛细管电压等参数,扫描质荷比范围 65~1500,采集并分析产物质谱图谱。
酶动力学参数测定
设置梯度底物浓度进行酶促反应,严格控制酶用量保证反应速率与蛋白浓度呈线性相关;采用双曲线回归模型拟合动力学数据,每组实验设置三次生物学重复,计算获得米氏常数、最大反应速率、催化效率等动力学指标。
天然蛋白分子量测定
利用凝胶过滤色谱法测定重组蛋白天然聚合状态,选用标准分子量蛋白标志物标定色谱柱洗脱体积,结合酶活检测确定目的蛋白洗脱位置,判定蛋白天然分子量;同时通过变性聚丙烯酰胺凝胶电泳测定蛋白亚基分子量。
酶学性质分析
配制不同 pH 梯度缓冲液,测定两种酶的最适反应 pH;在反应体系中分别添加不同二价金属离子,探究金属离子对酶活的影响;仅针对 Ph-CNL 设置梯度钾离子浓度,明确钾离子浓度与酶活性的关联,所有实验结果均取三次独立实验平均值。
RNA 干扰沉默载体构建与植株遗传转化
以古代稀芳樟醇合酶基因LIS 花瓣特异性启动子驱动,构建Ph-CNL 与Ph-4CL1 基因 RNA 干扰载体,构建流程如下: 首先扩增长度为 1038 bp 的LIS 启动子片段,插入 pART27 载体;再将 pRNA69 干扰表达盒上的多克隆位点替换为反向排布 的 Gateway 重组位点,将完整干扰片段连接至改造后 pART27 载体中LIS 启动子下游。
分别截取Ph-CNL 473 bp、Ph-4CL1 441 bp 的基因特异性保守片段,对应基因序列位置依次为 96~568 位、214~654 位;先将片段连入入门载体 pENTR/D-TOPO,再通过 LR 重组反应,定向插入载体中两组反向 Gateway 位点,形成反向重复沉默片段。
采用农杆菌 GV3101 菌株介导的叶盘转化法 侵染矮牵牛,获得转基因植株。利用 PCR 扩增卡那霉素抗性基因nptII 筛选阳性株系;生根成活的阳性植株移栽至温室栽培。对 T0、T1 代植株人工自交收种,将种子播种于含 200 mg/L 卡那霉素的 MS 培养基,鉴定后代植株抗性分离情况。
花香挥发性物质测定
采用闭环吸附法收集野生型、CNL 及4CL1 干扰株系的花瓣挥发性香气物质。米切尔矮牵牛属于夜间释香型花卉,统一于当晚 20:00 至次日 8:00 收集花香组分。 为消除昼夜节律带来的实验误差,固定在晚间 20:00 采摘开花后第 2 天的花冠组织,称取 1 g 样品用于测定体内内源代谢物;样品经二氯甲烷浸提、浓缩处理后,借助气相色谱 - 质谱联用技术完成组分定量分析。
有机酸与辅酶 A 酯类物质含量测定
统一在开花后第 2 天晚间 20:00 采集花冠样品,规避昼夜节律影响,参照已发表成熟实验方法,测定植株体内游离有机酸及各类羟基肉桂酰辅酶 A 酯的内源积累量。
矮牵牛花瓣过氧化物酶体分离
参照经典分离方法并稍作改良,4℃低温环境下从 10 g 开花后第 2 天新鲜花瓣中提取过氧化物酶体。 样品经预冷研磨缓冲液充分匀浆,纱布过滤去除组织残渣;先后经差速离心初步富集细胞器,再利用 Percoll 密度梯度离心进一步纯化,收集蔗糖垫层上方白色条带即为高纯度过氧化物酶体组分。 经重悬洗涤去除介质杂质后,将沉淀重悬于磷酸钾保存缓冲液中,采用布拉德福德法测定蛋白浓度。 通过测定特征标记酶活性评估细胞器纯度:检测过氧化物酶体过氧化氢酶、线粒体延胡索酸酶、细胞质醇脱氢酶活性,同时测定叶绿素含量判定叶绿体污染程度;最终纯化组分中线粒体污染活性不足 1%,且未检测到细胞质酶活性,纯度符合实验要求。
RNA 提取与实时荧光定量表达分析
利用植物总 RNA 提取试剂盒,分别提取矮牵牛叶片、不同花器官、不同花期花冠,以及昼夜节律不同时间点的花冠总 RNA;除特殊说明外,样品均在下午 15:00 采集。 经 DNase I 消化去除基因组 DNA 污染后,反转录合成 cDNA。 使用基因特异性引物开展实时荧光定量 PCR,以延伸因子EF1α 作为内参基因校正上样量,所有引物扩增效率均处于 90%~100%。 采用标准曲线法 完成基因绝对表达量定量,以纯化的质粒扩增片段制作标准品绘制标准曲线,换算得到目的基因转录本绝对含量;每组实验设置 3~4 组生物学重复。
辅酶 A 连接酶蛋白亚细胞定位
以携带 35S 强启动子的 p326-sGFP 载体为骨架,构建多种绿色荧光蛋白融合表达载体:
- N 端荧光融合载体 :将完整Ph-4CL1 、Ph-CNL 编码序列分别连接在 GFP 基因下游;同时构建缺失 C 端 PTS1 定位信号 ARL 三肽的Ph-CNL 截短融合载体。
- C 端荧光融合载体 :将目的基因全长序列连接在 GFP 基因上游。
所有载体构建过程均通过测序验证序列无误。 制备拟南芥原生质体并完成质粒转化,培养 16~20 h 后利用激光共聚焦显微镜观察荧光分布;依次单独采集线粒体红色探针荧光、GFP 绿色荧光、叶绿体自发荧光信号,规避荧光串色干扰,结合荧光分布特征判定蛋白亚细胞定位。 设置对应激发光波长与荧光收集波段,完成图像采集与定位分析。