 image-20240815151428804
image-20240815151428804
2024年8月14日沙特阿卜杜拉国王科技大学 Brande B. H. Wulff 和 Simon G. Krattinger团队在Natue发表Origin and evolution of the bread wheat D genome研究论文,通过研究粗山羊草(Aegilops tauschii,也被称为节节麦),即面包小麦D基因组的供体,建立并分析了广泛的基因组资源,揭示了面包小麦D基因组的复杂演化历史,并发现了潜在的遗传多样性。
研究人员对46个粗山羊草基因组进行分析并克隆一个抗病基因,在复杂的抗病位点上进行单倍型分析,区分等位基因和旁系同源基因拷贝。该研究还探讨了野生近缘种在作物改良中的潜力,为未来的作物育种提供了重要的参考。
小麦的复杂起源历程
普通小麦是AABBDD异源六倍体,其形成涉及3个原始祖先物种和2次天然杂交。普通小麦是由祖先野生的一粒小麦(乌拉尔图小麦,含AA基因组)与拟斯卑尔托山羊草(Aegilops speltoides ,含BB基因组)杂交形成四倍体野生二粒小麦(Triticum turgidum ,含有AABB基因组)。随后在大约1万年前节节麦(DD)和四倍体小麦( AABB)自发杂交进而产生了六倍体小麦,也就是我们现在所说的普通小麦。节节麦是小麦D基因组的二倍体祖先,是小麦的重要资源。目前为止,六倍体小麦及其亲缘种AA、DD、AABB和AABBDD的精细基因组序列图谱均已绘制完成,这为小麦的功能基因组学、比较基因组学和进化基因组学研究奠定了基础。
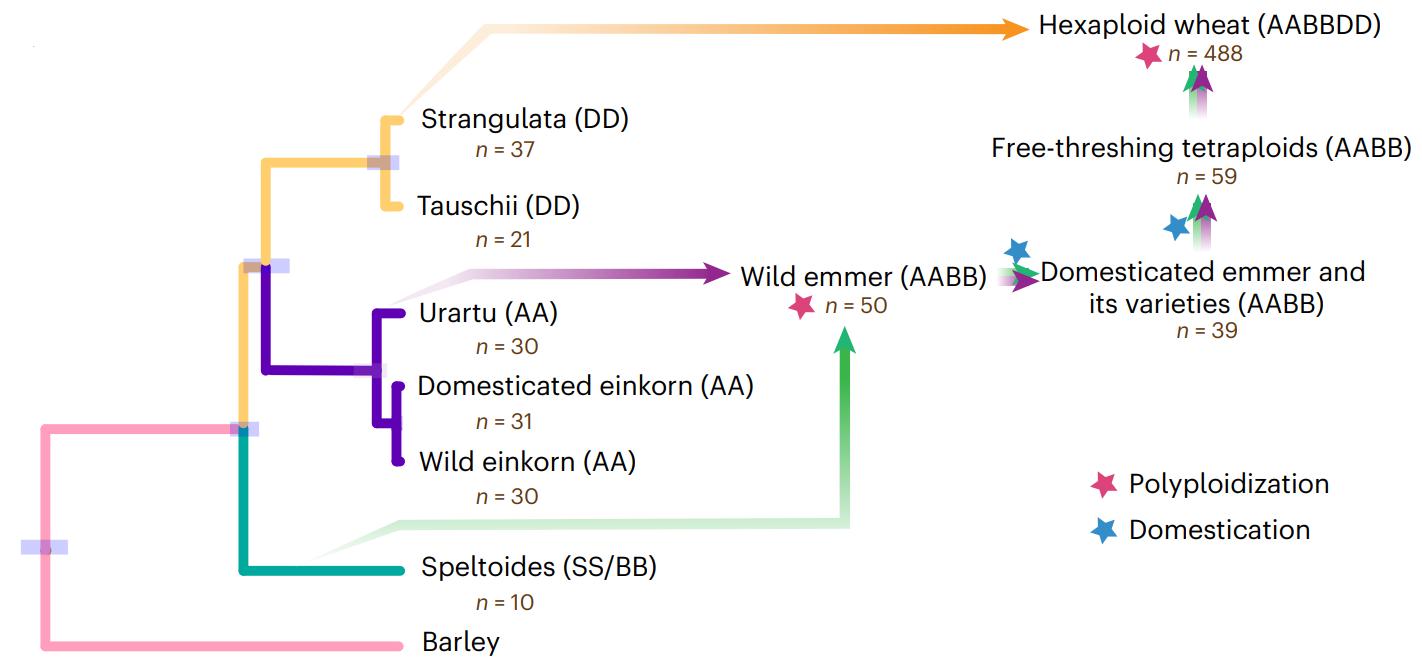 image-20240819200341214
image-20240819200341214
驯化过程中,面包小麦经历了多倍化(polyploidization),这显著减少了遗传多样性。然而,驯化后的反复杂交事件,即与野生小麦近缘种和其他驯化小麦品种的杂交,显著增加了面包小麦的遗传多样性。与野生祖先相比,现代面包小麦的遗传多样性较低,这是由多倍体化、驯化和育种瓶颈引起的,野生小麦的近缘种代表了基因库,并具有尚未被纳入面包小麦的多样性和有益等位基因。
与四倍体和二倍体祖先相比,D亚基因组的加入导致了面包小麦广泛的适应性和卓越的加工品质。因此,野生节节麦群体是小麦D亚基因组改良的种质资源库,找回小麦驯化过程中丢掉的基因,可以用来培育抗逆品种,或者获得一些现代小麦原来没有的特性,例如优良的抗病性。
节节麦基因组多样性资源
为了全面评估节节麦的遗传多样性,首先利用k-mer分析从920个已测序的节节麦种质中揭示了493个非冗余的节节麦种质,覆盖了从土耳其西北部到中国东部的地理范围,并定义了由3个基系划定的系统发育。
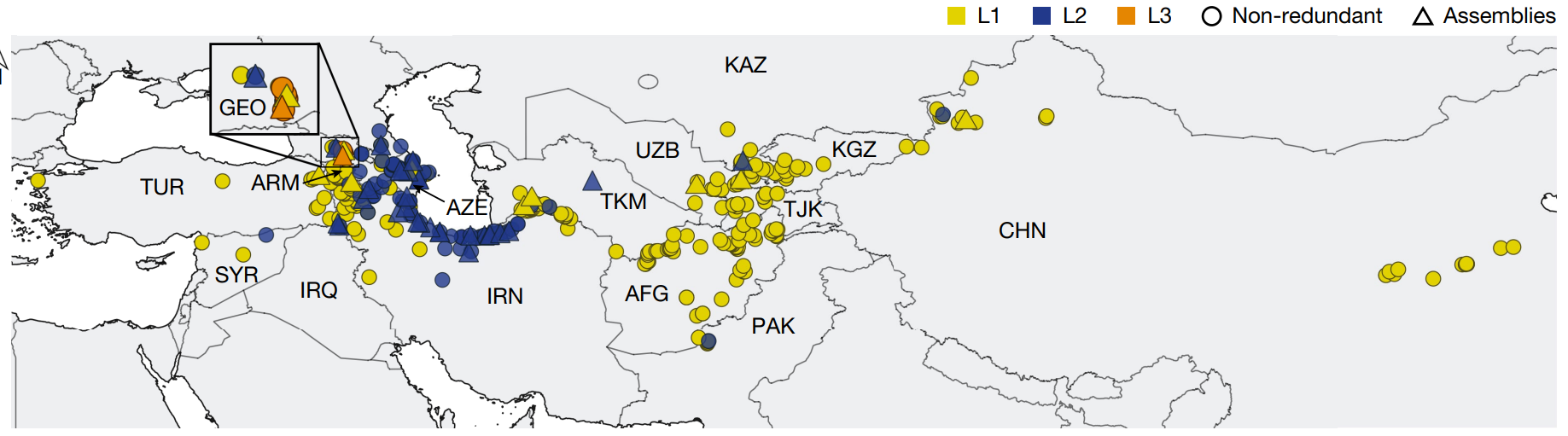 image-20240819202336163
image-20240819202336163
之后利用遗传多样性、地理多样性和表型多样性,基于K-mer分析筛选了46份种质,覆盖了72.5%的Tauschii多样性,包括11个L1种质、34个L2种质和1个L3种质。
通过 PacBio 测序对选定的 46 个种质进行了基因组测序,覆盖率中位数为 23 X,然后为每个谱系选择一个具有代表性的种质来生成从头注释的染色体尺度组装体。即 TA10171(L1)、TA1675(L2)和 TA2576(L3)。对于这三个种质,将测序覆盖率从18 至 47 倍增加到 67 至 97 倍,三个染色体尺度组装的从头注释显示了43,511至44,275个蛋白质编码基因。
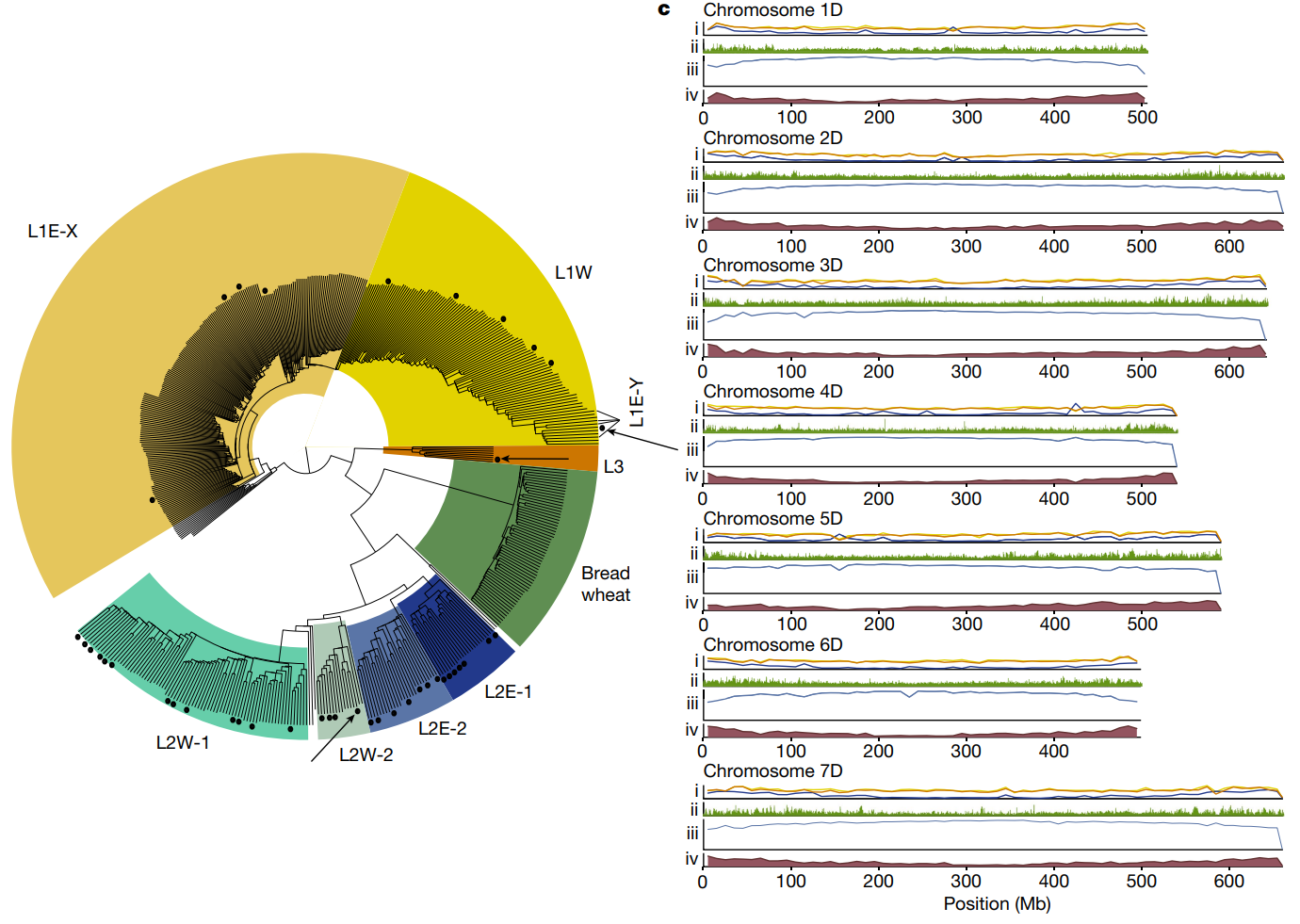 image-20240819202500126
image-20240819202500126
单倍型分析和抗病基因的克隆
为了评估节节麦基因组资源的价值,研究人员重点关注抗锈病相关基因,对 46 个节节麦基因组的分析后克隆了一个抗病基因,并进行了单倍型分析,从同源拷贝中辨别等位基因。染色体1DS内包含了秆锈病抗性基因SrTA1662 (重新命名为Sr66 ),通过对Ae. tauschii CPI 110799(Sr33供体)、AUS 18913、AUS 18911和TA1662(SrTA1662供体)中的抗性基因类似物(RGA)分析,反映出不同的Mla单倍型。
基于k-mer的全基因组关联研究(GWAS),利用Pt菌株BBBDB在Ae. tauschii TA1675组装中定位。染色体2D的峰值对应于抗叶锈病基因座LR39 。通过双亲定位LR39 区间,标记Csq21和Csq22以及与LR39 共分离的标记Csq8和Csq25-Csq30。然后通过VIGS进行验证,AL8/78是易感对照;BSMV-γGFP是表达GFP沉默构建体的大麦条纹花叶病毒(对照);BSMV-γLrSi2、BSMV-γLrSi6和BSMV-γLrSi7是针对WTK基因的特异性沉默构建体,BSMV-γLrSi3和BSMV-γLrSi8是针对NLR基因的特异性沉默构建体。作者进一步分析了Lr39 基因结构和Lr39蛋白结构,灰色框代表未翻译区,橙色框为外显子,线条为内含子。
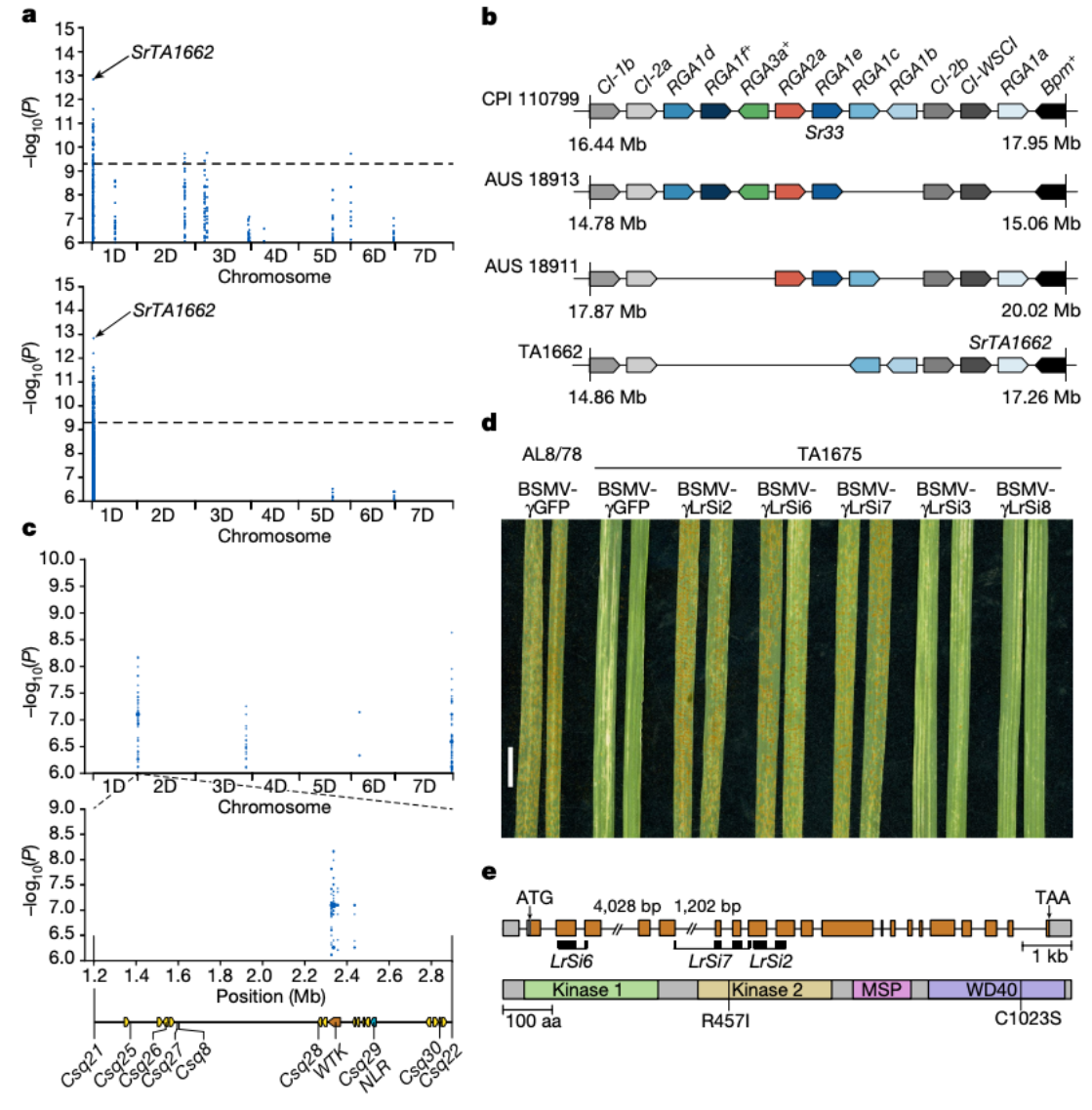 image-20240819202712688
image-20240819202712688
TA1675携带抗叶锈病基因Lr39,该基因定位于染色体2D短臂,与Pt分离物BBBDB(对Lr39无毒)29基于k-mer的关联结果显示,在染色体2DS的端粒末端有一个峰值信号,对应于TA1675组装中的2.33-2.45 Mb区域,该位置与Lr39侧的标记重叠,这些标记是根据双亲本遗传作图确定的。关联峰所在的基因组区域在TA1675中包含3个候选基因(1个小麦串联激酶(WTK)和2个功能未知的基因),通过双亲本定位鉴定的区间包含16个基因。在功能注释和多态性的基础上与易感材料进行比较,发现TA1675中WTK候选基因的病毒诱导基因沉默(VIGS)导致对叶锈病的易感性增强。
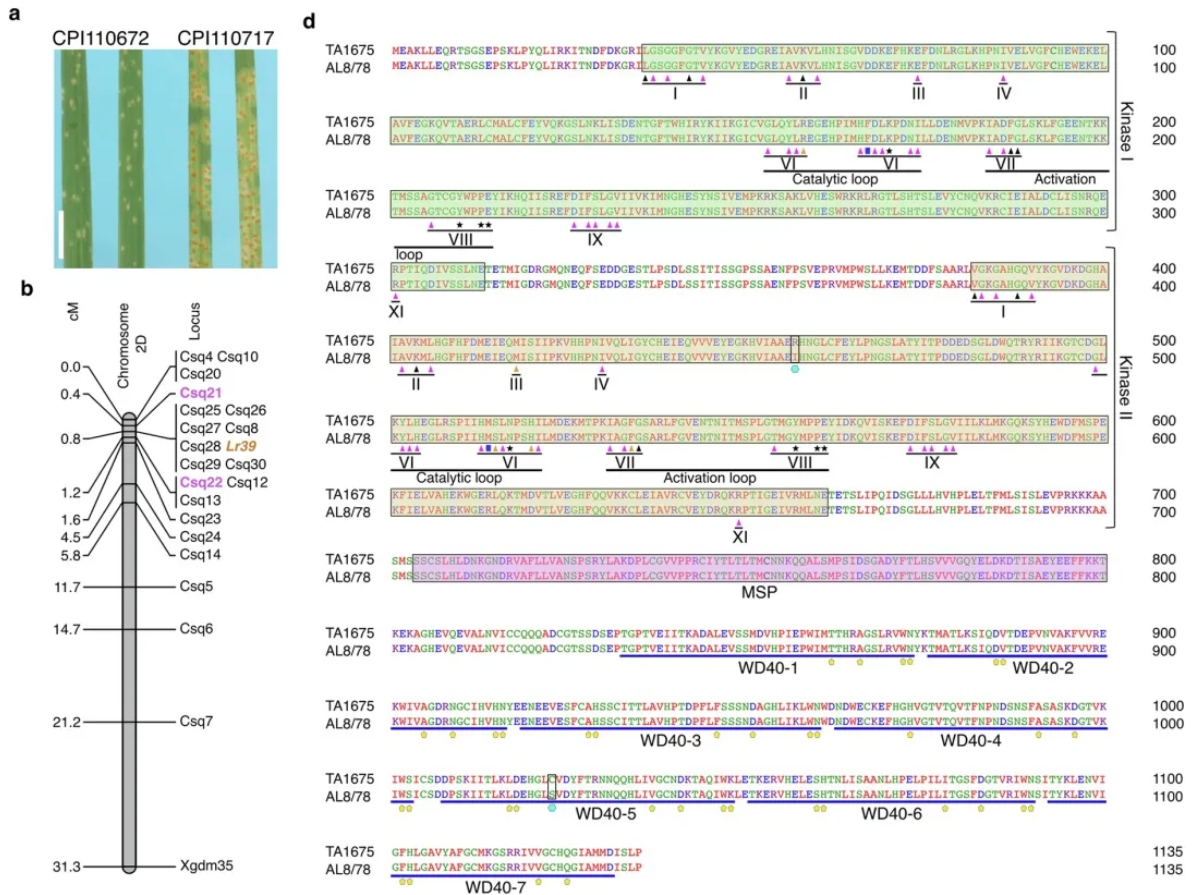 image-20240819213156493
image-20240819213156493
研究人员重点关注小麦抗锈病相关基因,包括小麦条锈病、小麦叶锈病、小麦秆锈病三类,根据前人的研究,秆锈病抗性基因SrTA1662 从节节麦种质TA1662渗入到面包小麦中,并定位于染色体1DS的秆锈病抗性位点SR33,由于原始定位无法确定TA1662的秆锈病抗性基因是新基因还是与Sr33等位基因,因此该基因被临时命名为SrTA166223 。Sr33和SrTA1662均编码属于Mla家族的细胞内核苷酸结合富亮氨酸重复序列(NLR)免疫受体。
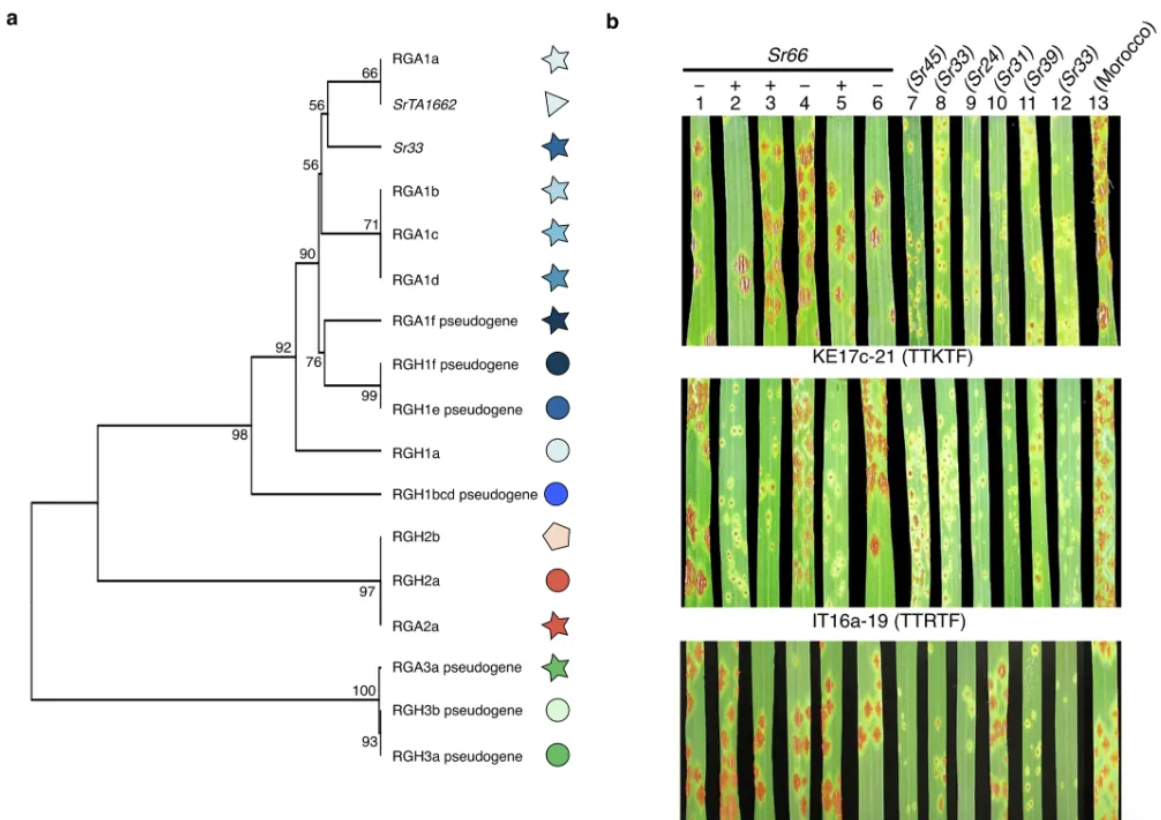 image-20240819212738219
image-20240819212738219
作者通过基于k-mer的关联分析,初步发现了SrTA1662 候选基因,详细的单倍型分析显示,SrTA1662 是Sr33 的旁系同源基因,而不是等位基因。比较仅携带Sr33 或SrTA1662 的Ae. tauschii 品系的秆锈病侵染表型时,观察到这两个基因似乎具有不同的特异性,作者通过接种SrTA1662 转基因小麦品系证实了这一观点。分析表明SrTA1662 对Sr33 上的Pgt分离株的一个子集产生了抗性,符合小麦基因命名标准,因此作者将SrTA1662 重命名为Sr66。
使用含有抗性基因的种质的参考基因组组装来进行基因克隆和命名很重要。如果使用以前的基因组,在这里进行的分析将会比较困难,因为它们都不是Sr33、SrTA1662 (Sr66)或Lr39的供体材料,也就是说参考基因组里可能压根没有想要克隆的目标基因。
小麦D基因组经历了更为复杂的长期驯化过程,研究人员通过分析发现反复出现杂交和基因流慎入,导致D基因组出现拼凑式单倍型。里海西南部的L2群体是面包小麦D基因组的主要供体,但是还携带了来自不同亚群的基因组片段。
小麦D基因组中约有1%来自L3,其遗传独特性和地理限制使得这一谱系成为研究基因渗入空间动态的理想例子,作者假设具有较高L3基因组含量的面包小麦地方品种,可能代表L3基因渗入的更古老的状态,为了鉴定L3基因组比例高于平均水平的面包小麦品种,研究人员开发了MissingLink Finder("缺失链接查找器")
缺失链接查找器估算一个种或系特定的参考k-mer集与从单个小麦材料的基因分型数据生成的样本k-mer集之间的相似性,并将结果计算为Jaccard相似性系数。作者使用了一个包含7.69亿的参考k-mer集,并将其与来自82,293个基因型小麦材料(每个616万k-mer)的单个样本k-mer集进行比较,鉴定出了503个面包小麦品种具有高于平均水平(超过两倍)的标准化Jaccard指数,表明Ae. tauschii L3含量增加,Jaccard指数最高的139份材料是六倍体小麦,作者还发现了364个面包小麦地方品种的L3基因渗入比例明显增加。
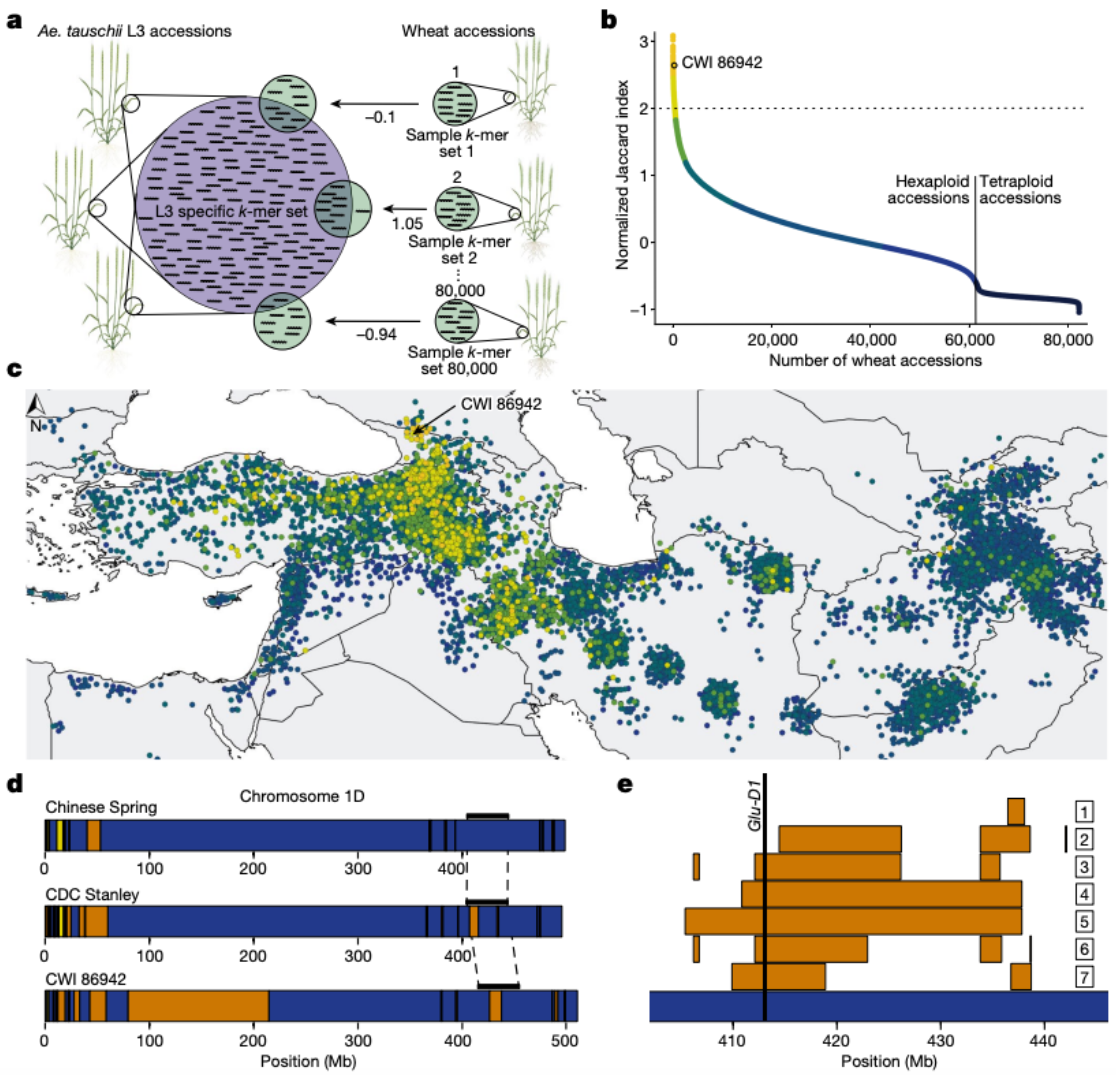 image-20240819203046682
image-20240819203046682
不同亚群对六倍体小麦的贡献
a图展示了小麦D基因组的各亚群比例。内圈用实色表示在17个六倍体小麦组装中的平均比例,外圈浅色表示17个小麦基因组中发现的最大比例,并基于代表性材料为每个亚群分配了地理位置,饼图展示了不同亚群对小麦D基因组的贡献比例。b图展示了面包小麦D基因组的最少杂交事件数量,染色体图展示了中国春小麦染色体1D、6D和7D上的单倍型块,颜色代表不同的节节麦亚群。
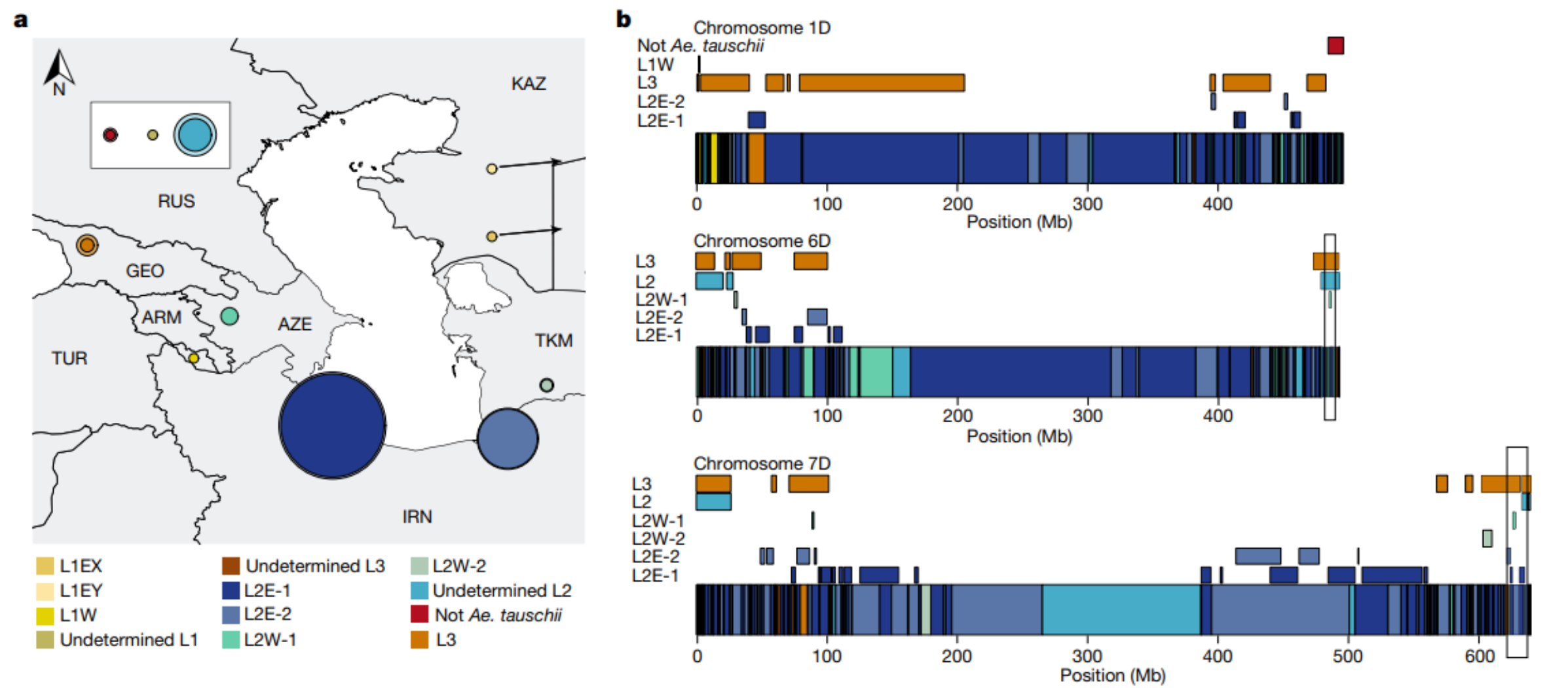 image-20240819204514280
image-20240819204514280
本文通过对Tausch的山羊草(Aegilops tauschii)的基因组资源进行深入分析,揭示了面包小麦D基因组的起源和演化过程。并建立了46个Ae. tauschii基因组的高质量基因组组装,并进行了单倍型分析,成功克隆了抗锈病基因。
小麦D基因组的遗传多样性不仅来自主要的L2亚群,还包含了L1和L3亚群以及其他不同地理来源的亚群的贡献,为利用野生小麦亲缘种的遗传资源进行作物改良提供了新的视角。野生种不仅能够帮助我们更好地理解小麦如何适应不断变化的环境条件,而且还能指导育种家们培育出更耐旱、抗病、高产优质的新品种。
原文链接:https://www.nature.com/articles/s41586-024-07808-z
本文由mdnice多平台发布