稳定细胞系平台的技术目标是在可传代的细胞群体中,使外源表达单元以可继承的基因组构型长期存在,并在多代培养中维持相对一致的转录活性与产物特征。与瞬时转染中质粒DNA在细胞内短期表达不同,稳定表达依赖"整合构型---选择压力---染色质状态"三类因素共同决定:整合构型影响可及性与拷贝组织,选择压力决定表达单元在群体中的保留与扩增趋势,染色质状态决定转录是否在传代中发生衰减。
一、 基因组整合机制与位置效应
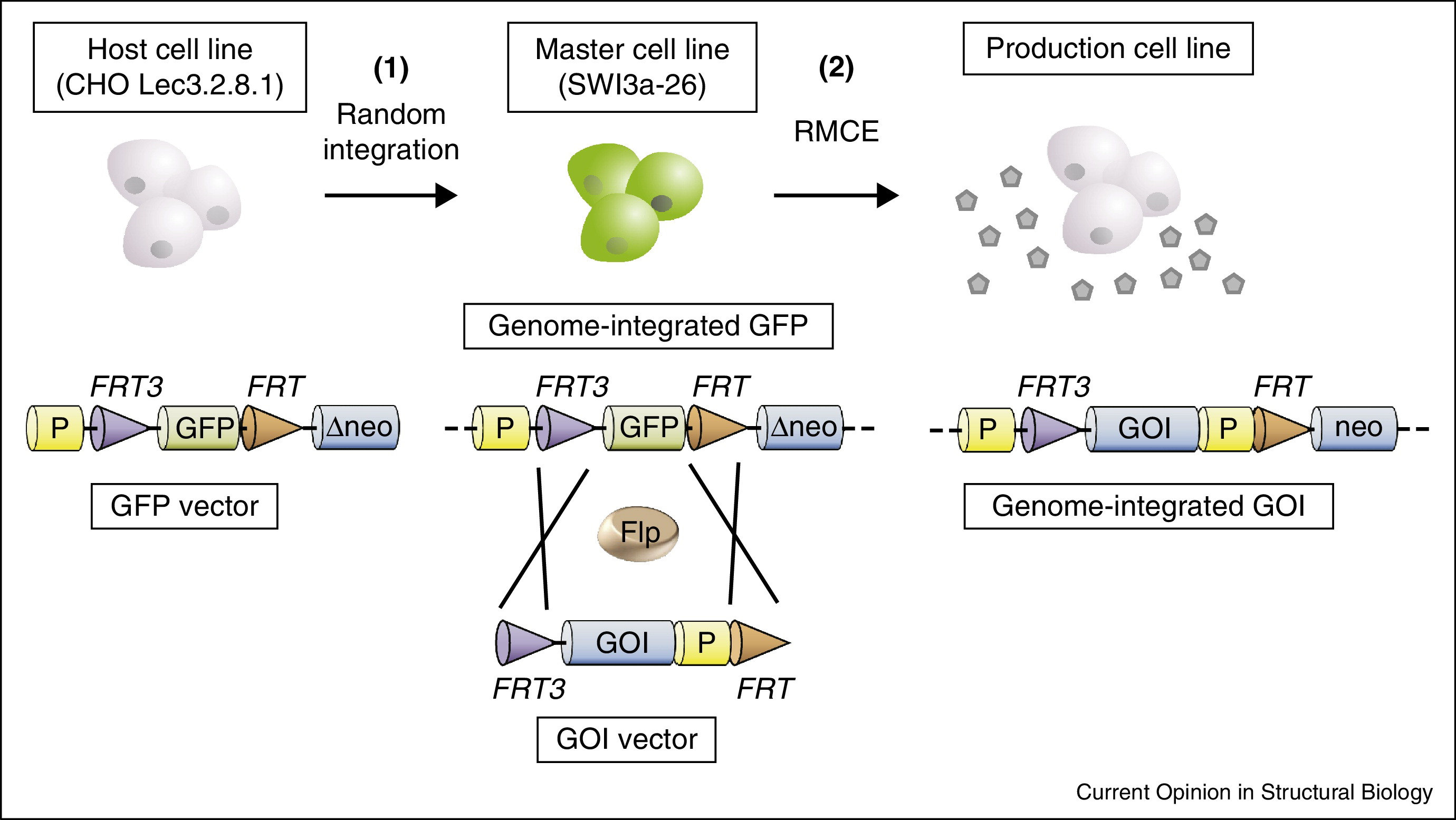
外源表达载体进入细胞核后,若形成稳定整合,通常与DNA损伤修复相关途径共同作用,其中非同源末端连接(NHEJ)是典型路径之一。细胞将外源DNA末端识别为需修复的断裂结构并完成连接,从而使表达盒与宿主染色体形成共价连接。由于该过程多为非定点事件,整合位点的染色质环境会显著影响转录可及性,即位置效应:整合落入异染色质区域时,启动子及增强子更易受到抑制性修饰影响,表现为转录活性偏低或在传代中出现沉默倾向;整合位于染色质更开放区域时,转录相关因子更易持续访问表达单元,表达水平与稳定性通常更有利。
二、 DHFR与GS选择体系对拷贝结构与表达负荷的约束
稳定细胞系平台常通过代谢选择体系将"表达单元存在"与"细胞生存"建立耦合关系,使携带并表达目标构建的细胞在群体中获得选择优势。DHFR体系与GS体系的共同点是:在特定营养或代谢约束下,细胞必须通过表达外源选择标记相关酶活性维持生存,从而提高表达单元被保留的概率,并在部分条件下伴随拷贝扩增现象。
需要强调的是,拷贝数增加并不必然线性转化为产物增加。拷贝扩增提升mRNA供给的同时,会显著抬升翻译、折叠、装配与分泌通路负荷,进而影响细胞生理状态与产物异质性分布。因此,平台评价通常应同时关注"表达通量"与"加工能力匹配":在CHO细胞或HEK293细胞等哺乳动物表达系统中,高产表型更接近于外源表达通量与宿主加工容量达到可持续平衡的状态,而不是单一由选择压力强度或拷贝数决定。
三、 表观遗传状态与长期转录持续性
稳定细胞株在连续传代中出现表达衰减,常与表观遗传调控相关。外源表达单元整合后,局部DNA甲基化、组蛋白修饰变化及染色质构象收缩可能降低启动子可及性,导致转录活性逐代下降。此类变化可能在拷贝结构未明显改变的情况下发生,因此需要将结构存在与功能可及区分讨论。
为降低沉默风险,表达构建中可引入有助于维持开放染色质状态的顺式元件(如UCOE、MAR等),其讨论重点在于:在长期培养中维持启动子区域的可及性与转录起始概率,使表达行为更接近可重复状态。对稳定性的判断更适合基于多代传代后的比较结果,而非短周期表达峰值。若要形成可复核结论,建议并行比较拷贝结构(qPCR等)、转录水平及蛋白层面指标,以避免将单一层面的变化过度归因。
四、 蛋白稳态约束与产物一致性的表征指标
高表达背景下,细胞的蛋白稳态面临更高负荷,内质网折叠、装配与分泌能力可能成为限制因素。稳态受扰动时,产物层面可表现为分子群体分布变化,例如聚集相关组分比例上升或片段化比例变化。对稳定细胞系平台而言,高表达应与产物一致性并行评估,否则在后续研究中容易出现批次或传代依赖突变。
五、 总结
稳定细胞系平台的核心机制可归纳为三类决定因素:其一,NHEJ等整合路径形成的基因组构型与位置效应决定转录可及性并造成克隆间差异;其二,DHFR/GS选择体系通过代谢约束提高表达单元保留概率,并在一定条件下影响拷贝结构与表达负荷;其三,表观遗传状态决定长期转录持续性并关联沉默风险。在此基础上,将qPCR等遗传层面指标等纳入评价,有助于对克隆稳定性与产物一致性建立更清晰的机制解释。