A dual-substrate methyltransferase catalyzes consecutive methylation reactions to convert proline into stachydrine in Leonurus japonicus
益母草中双底物甲基转移酶通过连续甲基化反应催化脯氨酸生成水苏碱

益母草(Leonurus japonicus)为我国传统大宗中药材,被收录于《中国药典》,是唇形科中极具特色的药用植物。唇形科多数物种主要以萜类化合物为核心活性成分,而益母草则特异性富集大量生物碱。其中,水苏碱与益母草碱是其主要特征性化学成分与药效物质基础,且益母草体内的水苏碱含量约为益母草碱的 10 倍。
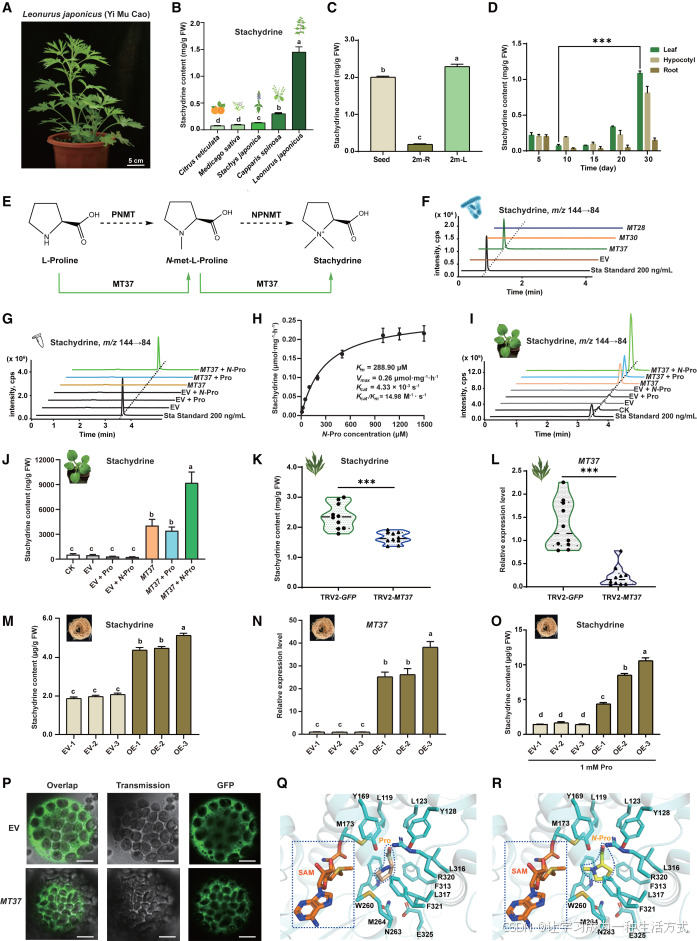
图1 益母草中水苏碱合成关键酶MT37的鉴定与功能解析
(A)益母草整株植株表型。比例尺,5 cm。
(B)多种已报道产水苏碱植物的水苏碱含量对比。数据为平均值±标准差,n=3个生物学重复。不同字母表示经单因素方差分析后存在显著差异(p < 0.05)。
(C)水苏碱在二月龄益母草种子、叶片和根系中的组织特异性分布。2m-L:二月龄植株叶片;2m-R:二月龄植株根系。数据为平均值±标准差,n=3个生物学重复。不同字母表示经单因素方差分析后存在显著差异(p < 0.05)。
(D)幼苗发育阶段(5--30 d)不同组织中水苏碱的动态积累特征。叶片水苏碱含量在第30天显著升高(n=3个生物学重复;非配对双尾t检验,***p < 0.001)。
(E)水苏碱合成通路预测(黑色虚线)与实验验证通路(绿色实线)。原有预测通路认为,L-脯氨酸经脯氨酸N-甲基转移酶(PNMT)催化生成N-甲基-L-脯氨酸,再由N-甲基脯氨酸N-甲基转移酶(NPNMT)催化生成水苏碱;而本研究证实,两步甲基化反应均由MT37单一酶催化完成。
(F)候选甲基转移酶的大肠杆菌体外酶活检测;仅MT37检测到显著催化活性(约220 ng/ml)。EV:空载体对照。
(G)纯化MT37蛋白的底物偏好性分析。Pro:脯氨酸;N-Pro:N-甲基脯氨酸。MT37可利用N-甲基脯氨酸为底物催化合成水苏碱,产量达570.63 ng/ml。数据为平均值±标准差,n=3个生物学重复。不同字母表示经单因素方差分析后存在显著差异(p < 0.05)。
(H)MT37蛋白的酶学动力学特征。
(I、J)MT37在本氏烟草中瞬时表达的体内功能验证,证实其可在植物体内催化脯氨酸合成水苏碱。数据为平均值±标准差,n=3个生物学重复。不同字母表示经单因素方差分析后存在显著差异(p < 0.05)。
(K)益母草中MT37病毒诱导基因沉默(VIGS)效果,沉默株系水苏碱含量显著降低(TRV2--GFP为对照)。非配对双尾t检验;***p < 0.001;n=10个生物学重复。
(L)qPCR结果表明,相较于TRV2--GFP对照,MT37沉默株系的MT37转录水平显著下调。非配对双尾t检验;***p < 0.001;n=10个生物学重复。
(M)益母草毛状根过表达MT37可显著提升水苏碱积累量。数据为平均值±标准差,n=3个生物学重复。不同字母表示经单因素方差分析后存在显著差异(p < 0.05)。
(N)MT37过表达毛状根株系中MT37的转录水平qPCR验证。数据为平均值±标准差,n=3个生物学重复。不同字母表示经单因素方差分析后存在显著差异(p < 0.05)。
(O)外源施加脯氨酸可进一步促进MT37过表达毛状根中水苏碱的积累。数据为平均值±标准差,n=3个生物学重复。不同字母表示经单因素方差分析后存在显著差异(p < 0.05)。
(P)MT37--GFP融合蛋白在本氏烟草原生质体中的亚细胞定位,定位于细胞质。比例尺,10 μm。
(Q、R)MT37分子对接模拟结果,脯氨酸与N-甲基脯氨酸均可稳定结合于MT37的同一活性口袋。蓝色虚线框代表甲基供体S-腺苷甲硫氨酸(SAM),蓝色不规则图形代表底物;图中标注了关键氨基酸残基及其位点。脯氨酸与N-甲基脯氨酸的氮原子距离SAM甲基碳的距离分别为4.7 Å和4.8 Å,符合该酶的催化机制特征。
水苏碱是一类结构简单、药理活性广泛的生物碱,广泛存在于微生物、植物及动物中,具有心脏保护、脑血管保护、抗纤维化和渗透保护等多种药理功能。在植物体内,水苏碱是重要的渗透调节物质,可通过调控胞内渗透压增强植株对盐胁迫、干旱胁迫的耐受性,且能在逆境条件下快速积累。同时,水苏碱可通过NodD2介导结瘤基因的表达调控,促进根瘤共生与固氮作用;还可通过降低幼虫存活率发挥抗虫作用,并调控黄酮类等防御代谢物的生物合成,参与植株代谢应答过程。多重生物学功能表明,水苏碱在植物生长发育与环境适应中具有重要的生理与生态意义。
尽管水苏碱分布广泛、生物学功能关键,但其生物合成通路在过去六十余年一直未被解析。早期紫花苜蓿研究证实脯氨酸是水苏碱的合成前体,但至今尚未鉴定出其合成关键酶。益母草可大量富集水苏碱,但其合成分子机制仍不明确,不同于该物种中已被解析的益母草碱合成通路。鉴于水苏碱重要的药用价值与广阔的应用前景,填补其合成机制的研究空白具有重要意义,也是本研究开展的核心动因。
本研究鉴定得到一种双底物甲基转移酶MT37,证实其为催化水苏碱生物合成的关键功能酶。综合利用转录组分析、体外酶活验证、大肠杆菌与本氏烟草异源表达、病毒诱导基因沉默、毛状根过表达及分子对接模拟等多重实验手段,系统完成了MT37的功能验证与水苏碱合成通路的解析,为水苏碱代谢工程改造提供了核心酶资源与关键分子理论依据。
代谢积累特征分析表明,益母草是水苏碱的高富集植物,其叶片中含量尤为丰富。组织分布结果显示,水苏碱主要积累于种子和叶片中,根系含量极低。发育动态分析发现,幼苗幼叶中水苏碱含量较低,在出苗20--30天期间含量急剧上升,表明水苏碱的生物合成受到严格的发育调控。
基于脯氨酸经两步连续甲基化生成水苏碱的预测通路,本研究结合转录组与代谢组关联分析,筛选得到42个候选甲基转移酶基因。通过聚类分析与系统发育分析进一步筛选候选基因,经大肠杆菌体外重组蛋白功能筛选,最终鉴定MT37可催化N-甲基脯氨酸转化为水苏碱。MT37的表达模式与水苏碱的积累规律高度吻合。体外酶活结果显示,纯化的MT37可高效催化N-甲基脯氨酸生成水苏碱(产量570.63 ng/ml,动力学参数KM=288.90 μM,kcat=4.33×10⁻³ s⁻¹),但无法直接利用脯氨酸作为底物。据此推测,MT37对脯氨酸的催化功能可能需要特定辅因子或胞内微环境的调控。由于植物体内环境更接近益母草原生环境,本研究进一步通过本氏烟草瞬时表达体系开展体内酶活验证。结果发现,MT37可在植物体内直接催化脯氨酸生成水苏碱,与原核表达体系的酶活特征存在显著差异,表明植物特有辅因子或翻译后修饰是介导第一步甲基化反应的关键。
为验证MT37在益母草体内的生物学功能,本研究利用烟草脆裂病毒介导的基因沉默体系开展体内功能验证。沉默MT37后,益母草叶片水苏碱含量显著下降,且MT37转录水平显著降低。因序列高度相似,部分株系中同源基因MT28的表达量出现轻微下调,但整体无显著统计学差异。与之相反,在益母草毛状根中过表达MT37可显著提升水苏碱积累量,qPCR结果证实其转录水平显著上调。外源施加脯氨酸可进一步促进水苏碱合成,表明底物供给是限制水苏碱合成效率的重要因素。
亚细胞定位结果显示,MT37定位于细胞质,与其在细胞质中催化氨基酸修饰的功能特征一致。蛋白同源建模与分子对接结果证实,MT37的活性口袋可同时容纳脯氨酸与N-甲基脯氨酸两种底物,从结构层面证实其为可催化两步连续甲基化反应的双底物甲基转移酶。
综上,本研究通过体外酶活、原核与植物异源表达、病毒基因沉默、毛状根过表达及结构模拟等多层次互补实验,全面、系统地验证了MT37的生物学功能。
本研究首次在植物中鉴定并解析了水苏碱合成关键酶的功能,填补了长期以来植物生物碱代谢领域的研究空白。水苏碱结构简单、物种分布广泛,但其合成通路一直未能被解析,主要受制于传统化学合成的研究局限、研究关注度不足以及关键中间产物N-甲基脯氨酸的不稳定性。本研究鉴定得到全新的脯氨酸N-甲基转移酶MT37,阐明了水苏碱合成的核心分子机制。植物体内实验证实,MT37可独立完成脯氨酸到水苏碱的两步连续甲基化反应;其中,脯氨酸向N-甲基脯氨酸的转化为限速步骤,依赖植物特有辅因子、蛋白翻译后修饰及胞内微环境。MT37具备典型的双底物催化活性,但两步甲基化反应为连续催化还是独立结合催化,仍有待进一步探究。此外,本研究在其他高含水苏碱物种中筛选得到序列相似度40%--50%的同源蛋白,表明MT37属于一类独特的新型N-甲基转移酶亚家族。
水苏碱物种分布广泛、兼具多种药理活性,在心脏保护、抗纤维化等领域具有重要应用价值。本研究阐明的水苏碱合成分子机制,为其开发利用提供了核心理论支撑。MT37可作为高效的合成生物学元件,用于定向合成水苏碱及甲基化氨基酸衍生物。后续可通过蛋白改造提升MT37的催化效率、拓宽底物谱、优化辅因子适配性,实现微生物或植物体系中水苏碱的规模化生物合成。该研究不仅为解析植物水苏碱的生理功能提供了新视角,也为其药用、保健产业化应用奠定了重要基础。