大家好,今天我们想和大家探讨植物界一个永恒的生存博弈------"生长与防御的权衡"。我们知道,氮素是植物生长的"油门",它驱动生物量积累;而干旱胁迫则是"刹车",它通过积累脱落酸来抑制生长以保命。在自然界复杂多变的环境中,植物常常同时面临着两个相互冲突的信号。那么,植物究竟是如何在"油门"和"刹车"之间做出生死抉择的?这是一个长期困扰我们的科学问题。
今天我们要解读的这篇PNAS文章《Antagonistic regulation of nitrogen and drought signaling mediated by NIN-like protein 7 transcription factor in Arabidopsis thaliana》首次揭示了一个关键的决策者------转录因子NLP7。研究发现NLP7作为汇聚转录回路的中心整合器,使植物能够通过调控氮和ABA/干旱信号通路来平衡生长和胁迫。

- 研究技术:ChIP-seq、RNA-seq、TARGET等
研究背景
植物需在资源有限的自然环境中权衡生长与生存,但二者信号通路常相互拮抗,分子整合机制尚不明确。尽管已知N(氮素)和WD(干旱,Water Deficit)分别通过NLP7(促生长)和ABA(促胁迫响应)通路调控基因表达,但二者交叉调控的转录网络核心因子未被系统揭示。先前研究多聚焦单一信号,缺乏多因素交互分析。
研究结果
1. 转录组学数据集的荟萃分析揭示了氮素、干旱与脱落酸(ABA)之间的交互作用
为了探究氮素与干旱的共性,作者并未局限于单一实验,而是进行了一项转录组元分析(Meta-analysis)。整合了37项公开研究,除了氮素(N)和干旱(WD)数据,研究特意引入了ABA处理的数据,因为ABA被认为是连接氮素与干旱信号的关键桥梁。样本涵盖了不同植株年龄、组织部位及多种实验条件。为确定响应基因,研究针对N、干旱和ABA各组实验分别鉴定了差异表达基因(DEGs)。设定了严格的统计阈值,定义了"共享差异表达基因"(sDEGs)。这些基因中约有10%是转录因子,筛选出的氮素响应基因与过往文献及功能注释高度吻合,证明了方法的可靠性。
此外,研究发现氮素信号与胁迫信号在转录层面存在惊人的重叠:约25%的干旱/ABA响应基因同时也出现在氮素(N)基因集中。超过50%的氮素(N)相关基因同时也受到干旱或ABA的调节。这种高比例的基因重叠直接证明了氮素信号与干旱/ABA信号之间存在紧密的功能耦合,为解析两者平衡机制提供了坚实的数据支撑。
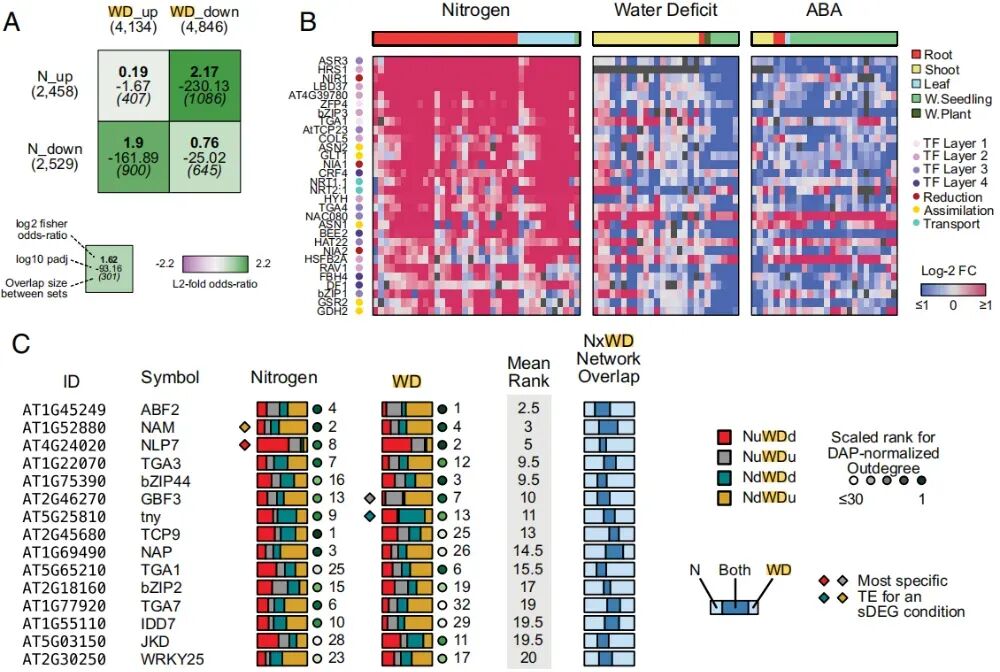
图1:转录因子介导拟南芥氮和干旱间的拮抗基因响应
2. 氮与干旱表现出拮抗基因响应
通过对多维转录组数据的深度挖掘,研究团队在全基因组水平确立了氮素与干旱/ABA信号之间显著的"反向拮抗"关系,即氮素激活的生长基因会被干旱/ABA强力抑制。进一步分析发现,高达82%的共调控基因受ABA主导,且这种拮抗模式在根与地上部中高度一致。这有力地证实了植物在干旱下的生长抑制,并非源于物理上的养分阻断,而是通过ABA介导的主动信号对话,在全株范围内系统性地关闭了氮素响应通路,从而实现从生长到防御的策略切换。
3. 常见转录因子调控氮素与干旱响应
作者利用GENIE算法,分别构建了氮素(N)和干旱(WD)的基因调控网络。研究探究了哪些转录因子作为氮(N)和干旱响应的中枢调控因子。根据每个转录因子对响应的特异性为其分配了等级,然后进行特异性评分,筛选出了前15名候选TF。NLP7作为众所周知的氮素主效调控因子,在氮素网络中排名第8。但在干旱网络中,竟然高居第2位!更重要的是,NLP7被预测为NuWDd模式(即氮素上调、干旱下调)的最特异性调控因子(图1C)。这一计算分析精准地将NLP7 指向了"生长-胁迫平衡"的操纵杆位置,它不只是管氮素,更是决定植物在干旱下是否要关闭生长基因的关键开关。
4. NLP7是氮素与干旱相互作用的重要调控因子
已有报道显示nlp7突变体植物表现出对WD耐受的表型。然而,NLP7在WD条件下(无论是否存在N)对基因表达的具体影响尚未得到充分研究。为此,研究设计了一个包含三个维度的转录组实验:(1)基因型 (G):野生型 (Col-0) vs. nlp7突变体;(2)氮素水平 (N):高氮 (High N) vs. 低氮 (Low N);(3)干旱进程 (WD):第0天 (对照) vs. 第6天 (干旱)。采用三因素方差分析,结果显示,单一因素不重要,仅受单一因素(如只受氮、或只受干旱)影响的基因极少(不足5%)。约95%的差异表达基因都涉及两个或以上因素的交互,说明这三个系统是深度耦合的。
两大核心交互群,G:WD(基因型×干旱,约1700个基因):这组基因的变化依赖于NLP7,但与氮素浓度无关。这说明NLP7本身就是一个独立的干旱调节因子。G:N:WD(基因型×氮素×干旱,约850个基因):这组基因最复杂,它们同时受NLP7、氮素和干旱三者的共同调控。这正是研究要找的"生长-胁迫平衡"的核心基因集。作者进一步分析了这些受NLP7操控的基因到底在干什么。GO富集分析显示被干旱抑制的基因 (WD-repressed in G:N:WD)主要涉及光合作用、叶绿素合成、氮素代谢;被干旱激活的基因主要涉及抗旱响应基因(如RD21A、SnRK2.2、PP2CA)。
综上所述,这些结果表明NLP7协调调控着那些对氮(N)和干旱(WD)条件均有响应的基因,以及那些仅对干旱有响应的基因。这些基因参与和光、氮及干旱适应性相关的生物学功能。
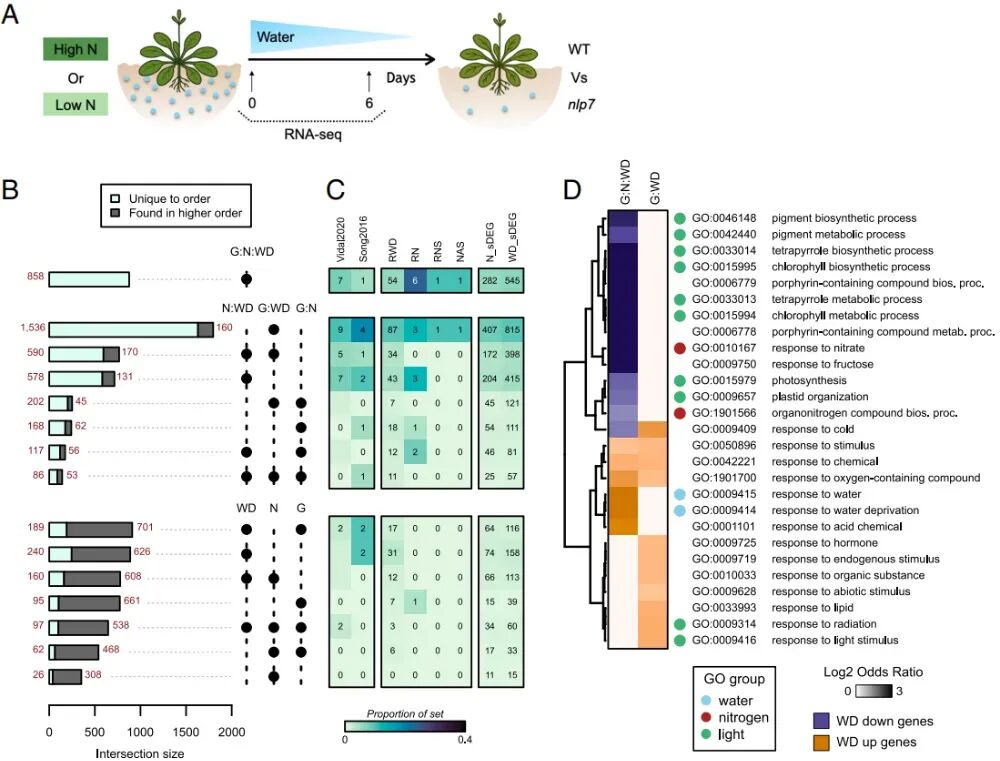
图2:NLP7在干旱条件下调控基因表达,并协调氮素与干旱之间的相互作用。
5. NLP7通过直接调控次级ABA及干旱相关转录因子整合氮素与干旱信号
研究使用了带GFP标签的NLP7转基因植株,分别在正常浇水(WW)和干旱处理(WD)6天后提取叶片,进行ChIP-seq实验。NLP7主要结合在基因的转录起始位点(TSS)和终止位点(TTS)附近,符合经典转录因子的特征。正常浇水(WW):NLP7结合了约1200个基因。干旱处理(WD):NLP7 结合了约5300个基因。其中4100个基因是NLP7仅在干旱条件下才去结合的,这暗示NLP7的DNA结合能力受水分可用性显著影响。
研究将ChIP-seq与之前的Meta-analysis进行比对,发现NLP7结合的基因中,显著富集了那些通常被ABA或干旱(WD)激活的基因,特别是转录因子。为验证NLP7是否直接诱导或抑制这些ABA或WD上调基因,研究使用了TARGET实验。实验结果显示,NLP7直接抑制了一批关键的ABA/干旱信号转录因子,包括HB6、ABF4、HAT22等著名的ABA信号通路基因。
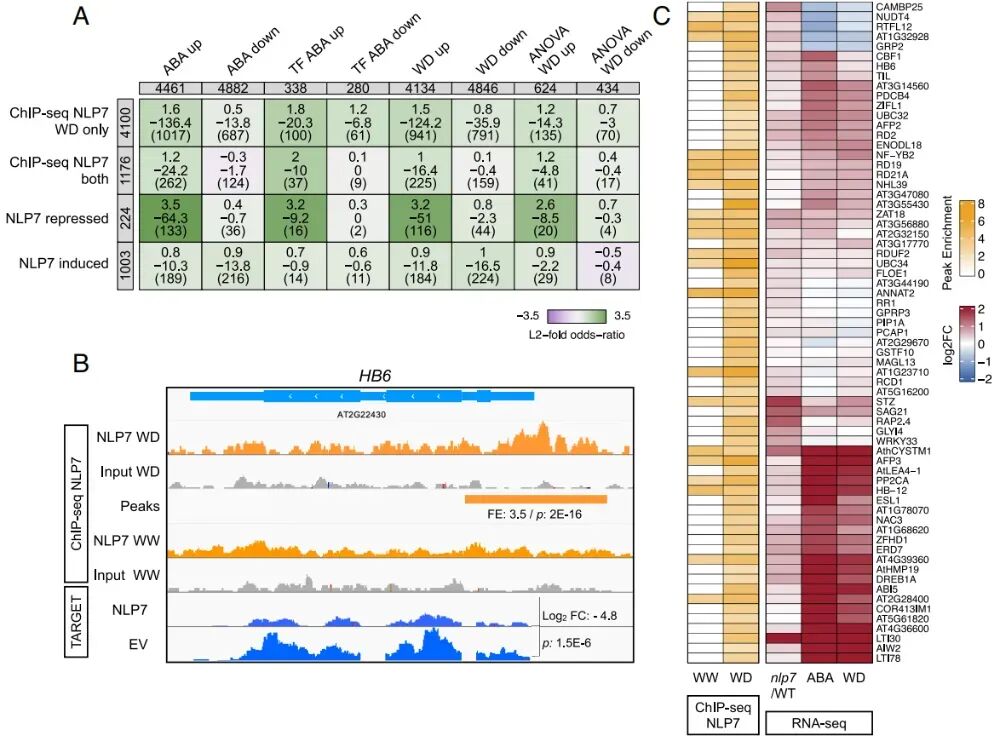
图3:NLP7直接抑制HB6及其他ABA/干旱响应转录因子
为进一步探究NLP7在N与WD相互作用中的转录调控机制,研究将生成的数据整合至单一基因调控网络(GRN)模型中。研究构建了一个包含直接结合(ChIP)、直接调控(TARGET)以及功能注释(GO)的综合网络,NLP7位于金字塔顶端。多个在ABA信号传导和WD耐受性中具有明确功能的转录因子被NLP7结合并直接抑制,包括HB6、NAC6、NAC083、WRKY18和NAP。利用DAP-seq数据,研究计算了NLP7对抗旱基因(RWD genes)的控制方式,发现NLP7直接靶向529个RWD基因,间接靶向508个RWD基因。
研究利用DAP-seq数据进一步探究不同次级转录因子如何调控G:WD和G:N: GWD基因集中的NLP7依赖基因。WRKY18和HB6的下游靶标主要富集在"纯干旱响应"基因集(G:WD)。这意味着NLP7通过控制它们来调节基础的抗旱反应。TGA7的下游靶标则主要富集在"氮-旱互作"基因集(G:N:WD)。这意味着TGA7是处理营养与胁迫交叉信息的关键节点。
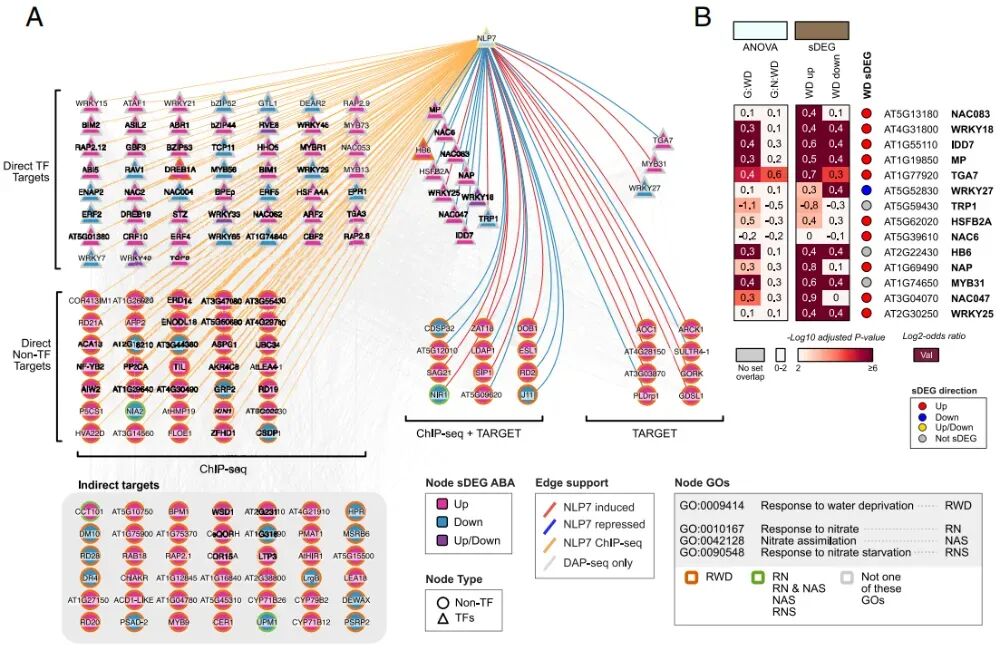
图4:NLP7直接和间接调控氮-水亏缺相互作用的转录网络
6. NLP7通过水分滞留和ABA介导的气孔关闭调节干旱耐受性
研究通过表型分析进一步阐明nlp7突变体对WD的响应特征。在干旱致死实验中,野生型的存活率仅为 60%,而nlp7突变体的存活率接近100%。为探究nlp7突变体的耐旱表型是否可与NLP7对植物生长的影响解耦,研究评估了在补充谷氨酰胺条件下的存活情况。结果表明,nlp7的耐低氮表型并非单纯由硝酸盐条件下的生长受损所致,而是与NLP7依赖的ABA信号通路破坏相关。
为深入解析NLP7基因在胁迫响应中的生理功能,研究分别对Col-0、nlp7及35S::NLP7植株进行胁迫试验。在同等水分供应条件下(SWC为20%),nlp7突变体的 相对含水量(RWC) 高于Col-0植株,而35S::NLP7植株则表现出较低的RWC。这些结果表明,nlp7植株增强的水分保持能力并非仅源于生长抑制,而是反映了对干旱(WD)的特异性生理适应。
为深入探究这一机制,研究采用ABA响应性检测法对气孔反应性进行了评估。ABA处理后,与Col-0植株相比,nlp7植株叶片气孔开度减小,而35S::NLP7品系则保持较宽的气孔开度,表明其ABA敏感性降低。这些结果共同支持以下模型:NLP7通过调控ABA介导的气孔调节和水分保存来影响水分亏缺耐受性,这凸显了NLP7作为生长与胁迫适应分子整合器的关键作用。
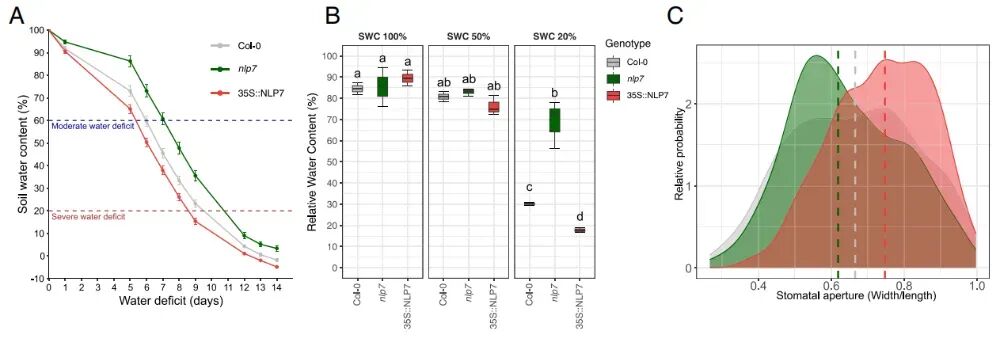
图5:NLP7调控植物水分保持及ABA依赖性气孔关闭。
这篇PNAS为我们提供了一个教科书级的"转录因子机制解析范式":
-
大数据指路:利用Meta-analysis (RNA-seq) 从海量数据中抽丝剥茧,锁定氮素与干旱的共调控基因集,不仅发现了"反向拮抗"的规律,更利用算法精准预测出核心操纵者------NLP7。
-
实锤验证:这是冲击高分文章最关键的一步。作者没有停留在转录水平的相关性分析,而是通过 ChIP-seq 技术,直接捕获了NLP7在全基因组上的结合位点。
-
网络构建:结合DAP-seq和TARGET实验,构建了层级分明的 GRN(基因调控网络),清晰地描绘出 NLP7 → HB6/NAC →气孔关闭/生长抑制 的完整信号级联。
给我们的科研启示:很多老师和同学的课题往往卡在这样的瓶颈:"手里有很好的表型(突变体抗旱/耐盐),转录组也测了一大堆差异基因,但就是找不到那个关键开关,讲不清直接机制。"这篇PNAS告诉我们:想发好文章,必须从"相关性"走向"直接调控"。
如果你想找全基因组的直接靶点,ChIP-seq或CUT&Tag都是不错的选择;
如果是难以制备抗体的非模式物种转录因子,DAP-seq则是更适合的选择;
如果你想看染色质的开放状态如何影响转录因子的结合,ATAC-seq 更是必不可少的利器。
在植物逆境与生长发育的研究中,表观遗传与转录调控永远是解释机制的最优解。如果您也想复刻这种由"表型"到"核心转录因子"再到"直接下游网络"的高分逻辑,欢迎联系我们。我们提供从ChIP-seq、DAP-seq到多组学联合分析的一站式服务,助您的课题从"现象描述"跃升为"机制解析",冲击属于您的Top期刊!