A minimal transcription factor network is sufficient to drive paclitaxel biosynthesis
一套最简转录因子网络即可驱动紫杉醇生物合成
紫杉醇的合成生物学技术路线综述--文献精读195_a synthetic biology roadmap for sustainable produc-CSDN博客
紫杉醇的药物代谢-文献精读73_紫杉醇球棍模型-CSDN博客

摘要
抗癌药物紫杉醇天然含量低、化学结构复杂,其供应一直受到限制。本团队此前已建立红豆杉形成层分生细胞(CMC)培养体系,研究发现茉莉酸甲酯可显著提升紫杉醇产量。本研究通过系统性分析,筛选出调控紫杉醇生物合成的核心转录因子。结果表明,两个 MYB 转录因子协同表达 ,可在稳定转化的形成层分生细胞中激活部分紫杉醇合成通路基因;最终使紫杉醇产量提升 121 倍,其重要前体物质巴卡亭 Ⅲ 产量提升 364 倍。本研究为高效生产紫杉醇提供了可行策略,该方法也可推广应用于其他高价值天然产物的合成优化。
引言
紫杉醇(商品名:泰素)是一种二萜类天然产物,也是全球重磅抗癌药物。1992 年,美国食品药品监督管理局(FDA)首次批准其用于卵巢癌治疗,此后其适应症逐步拓展至乳腺癌、胰腺癌及非小细胞肺癌等领域(1,2)。紫杉醇强效的抗肿瘤活性源于其复杂的分子结构,该物质以牻牛儿基牻牛儿基焦磷酸为起始底物,经由包含19 步以上酶促反应 的复杂生物合成通路生成(3-5)。目前,商业化紫杉醇主要依靠两种方式生产:一是半合成工艺,该工艺成本高昂且资源消耗大,需从红豆杉植株中提取巴卡亭 Ⅲ 等中间产物作为原料;二是依托生物反应器开展植物细胞培养(3,6)。紫杉醇被世界卫生组织列为基本药物,全球市场需求持续供不应求,因此亟需开发更具可持续性、可规模化的生产技术。
细胞工程培养体系是一种环境友好型紫杉醇生产技术,应用前景广阔(7-9)。目前,大肠杆菌、酵母等微生物体系已可合成紫杉醇早期前体物质(10,11),但由于难以高效表达细胞色素 P450 酶,该类体系的应用效果受到明显限制(12)。与之相比,植物细胞体系具备天然的氧化还原配对蛋白与亚细胞结构,更适合表达依赖细胞色素 P450 的代谢通路(13,14)。近年来,研究人员陆续鉴定出紫杉醇合成通路中多个关键缺失酶,包括 2'α- 羟化酶 T2′OGD、3'-N - 苯甲酰转移酶 T3′NBT(4)、紫杉烷环氧合酶 1(15)以及紫杉烷氧化辅助蛋白 FoTO1(5)。依托上述成果,研究人员已在异源宿主中成功重构出合成巴卡亭 Ⅲ 乃至紫杉醇的完整通路(11,12,15-18)。多个研究团队结合基因组学与合成生物学技术优化基因表达,在本氏烟草中实现了关键中间产物巴卡亭 Ⅲ 的异源合成,产量范围约为 50 纳克 / 克至 10~30 微克 / 克(5,15,16,18)。另有研究表明,在本氏烟草中表达最简基因组合后,巴卡亭 Ⅲ 产量已接近红豆杉针叶中的天然含量(18)。尽管上述异源表达体系为紫杉醇可持续生产提供了新思路,但紫杉醇与巴卡亭 Ⅲ 的产量仍偏低,难以满足工业化生产要求(19)。
本团队此前建立了源自植物维管干细胞的 ** 红豆杉形成层分生细胞(CMC)** 培养体系,并证实茉莉酸甲酯(MeJA)诱导可显著提升该体系中紫杉醇的合成量。本研究旨在挖掘紫杉醇生物合成的核心调控因子,进一步提高该细胞体系的紫杉醇产量。本研究鉴定并验证了两个 MYB 转录因子,二者可显著促进红豆杉形成层分生细胞合成紫杉醇。在稳定转化这两个 MYB 基因的红豆杉细胞株中,紫杉醇产量可达 98 微克 / 克鲜重,较对照组提升约 121 倍;关键中间产物巴卡亭 Ⅲ 产量达 877 微克 / 克鲜重,为对照组的 364 倍。本研究建立的转录因子改造策略,不仅揭示了这条复杂生物合成通路的层级调控机制,也为该产物的工业化转化提供了具备潜力的产量水平。
结果
筛选茉莉酸甲酯诱导后上调表达的转录因子
已有研究证实,茉莉酸甲酯可诱导东北红豆杉形成层分生细胞发生转录级联反应,同时显著促进紫杉醇合成(20,21)。本研究利用茉莉酸甲酯处理红豆杉形成层分生细胞,结果显示细胞内紫杉醇与巴卡亭 Ⅲ 含量均显著上升(附图 S1A),与前人研究结论一致(20,21)。
转录组分析结果表明,茉莉酸甲酯处理后,细胞基因表达呈现两次明显的表达波动:诱导 0.5 小时 时,大量基因率先上调表达;诱导 12 小时 时,基因表达再次发生变化,上调、下调基因占比分别为 40%、60%(图 1A)。经统计,茉莉酸甲酯处理后共鉴定到 867 个上调基因、749 个下调基因(附图 S1B、S1C)。
对差异表达基因开展基因本体(GO)功能富集分析,结果显示,差异基因在紫杉醇生物合成过程 、各类紫杉烷合成相关酶活性条目上显著富集,其中包括 10 - 去乙酰巴卡亭 Ⅲ-10-O - 乙酰转移酶等关键酶基因(图 1B、附图 S1D)。该结果充分说明,茉莉酸甲酯可激活紫杉醇生物合成的核心分子调控网络。
基因火山图进一步证实,诱导 2 小时后,紫杉醇合成通路基因发生大范围、极显著的上调表达(图 1C)。其中,编码通路前期、后期关键酶的基因(附图 S1E)表达上调尤为突出,包括紫杉二烯合酶(TASY)、紫杉二烯 - 13α- 羟化酶(T13αH)、紫杉烷 - 10β- 羟化酶(T10βH)、紫杉烷 - 2α- 醇 - O - 苯甲酰转移酶(DBBT)。以上结果明确证明,茉莉酸甲酯可直接激活紫杉醇合成所需核心酶基因的表达。
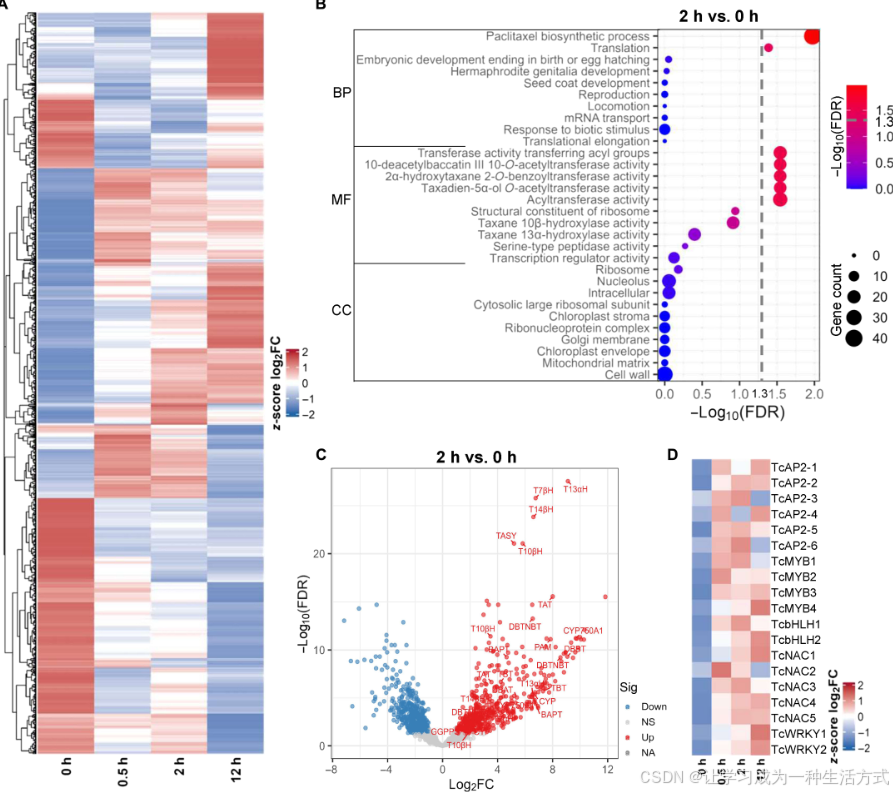
图 1 茉莉酸甲酯处理后形成层分生细胞(CMC)的转录组变化,筛选候选转录因子
(A) 红豆杉形成层分生细胞经 100 μmol/L 茉莉酸甲酯(MeJA)分别处理 0.5、2、12 小时后的基因表达热图。图中数值为经 Z-score 标准化的对数二倍变化值(log2FC) ,仅展示显著差异表达基因;行代表基因,列代表不同处理时间点。颜色由蓝至红依次表示基因表达下调、上调。 (B) 对比 0 小时与处理 2 小时样本,差异表达基因在 ** 生物学过程(BP)、分子功能(MF)、细胞组分(CC)** 三大基因本体(GO)分类中富集程度排名前十的散点图。圆点大小代表对应分类下的差异基因数量,颜色深浅代表统计学显著性[基于−log₁₀错误发现率(FDR)]。虚线代表−log₁₀ FDR = 1.3(即 FDR = 0.05)。 (C) 茉莉酸甲酯处理 2 小时后细胞的基因表达火山图(筛选标准:|log2FC| ≥ 1、FDR < 0.05)。红色:表达上调基因;蓝色:表达下调基因;白色:无显著差异;灰色:数据缺失。图中标注了紫杉醇合成通路中关键酶的编码基因。差异表达分析采用 DESeq2 软件完成,生物学重复数 n=3。 (D) 候选转录因子在茉莉酸甲酯不同处理时间点(0.5、2、12 h)的表达模式热图。
在总计 1646 个差异表达基因中,本研究鉴定出79 个转录因子 ,分属 19 个基因家族(附图 S2A)。其中 AP2/ERF(APETALA2)家族成员数量最多,其次依次为 MYB、碱性螺旋 - 环 - 螺旋(bHLH)、C3H 锌指蛋白及 NAC 家族。分析结果显示,茉莉酸甲酯处理可诱导其中 50 个转录因子上调表达,且上调基因主要集中在 MYB、C3H、NAC、WRKY 家族;另有 29 个转录因子表达下调,以 AP2、碱性亮氨酸拉链(bZIP)、CCAAT 类转录因子为主(附图 S2A)。
为筛选紫杉醇生物合成的正向调控因子 ,本研究重点关注茉莉酸甲酯处理 0.5 小时即出现显著快速上调 的转录因子,最终得到 19 个候选基因,包括 6 个 AP2、4 个 MYB、5 个 NAC、2 个 bHLH 及 2 个 WRKY 家族蛋白(图 1D)。这些转录因子的表达时序存在明显差异,暗示它们在转录调控中可能发挥不同功能(图 1D、附图 S2B)。例如,TcAP2-5 与TcMYB3 在处理 0.5 小时即被强烈诱导;而TcMYB4 、TcNAC1 、TcWRKY1 、TcWRKY2 则在 12 小时达到表达峰值(图 1D、附图 S2B)。 该表达时序特征表明:TcMYB3 可能参与紫杉醇合成基因早期第一轮转录激活 ,而TcMYB4 与 WRKY 转录因子则参与后续转录调控,负责精细调控或维持通路表达水平。研究进一步通过逆转录聚合酶链反应(RT-PCR)验证了上述转录因子受茉莉酸甲酯诱导上调的表达模式(附图 S2C)。
筛选茉莉酸甲酯诱导的转录因子,验证其对紫杉醇合成关键基因的调控作用
为探究候选转录因子对紫杉醇生物合成的调控能力,首先利用植物顺式作用元件分析工具 开展生物信息学预测(22),分析 10 个紫杉醇合成关键基因的启动子区域,包括:紫杉二烯合酶(TASY )、紫杉二烯 - 5α- 羟化酶(T5αH )、紫杉二烯 - 13α- 羟化酶(T13αH )、紫杉二烯 - 5α- 醇 - O - 乙酰转移酶(TDAT )、紫杉烷 - 10β- 羟化酶(T10βH )、紫杉烷 - 2α- 醇 - O - 苯甲酰转移酶(DBBT )、10 - 去乙酰巴卡亭 Ⅲ-10-O - 乙酰转移酶(DBAT )、苯丙氨酸变位酶、巴卡亭 Ⅲ-3 - 氨基 - 13 - 苯丙酰转移酶(BAPT )及 N - 苯甲酰转移酶(DBTNBT )(附图 S1E)。
预测结果显示,所有紫杉醇合成基因的启动子序列中,均含有一个或多个对应候选转录因子的特异性结合位点 ,且这类顺式元件在启动子区呈显著富集(附图 S3A、S3B)。
为实验验证转录因子与紫杉醇合成基因启动子的互作关系,本研究采用拟南芥高通量瞬时表达体系 结合双荧光素酶报告系统 开展体外验证(附图 S3C)。对 19 个候选转录因子逐一进行检测,分析其对上述 10 个紫杉醇合成基因启动子的转录调控活性。
经系统性筛选,共鉴定到 36 组显著的互作关系;其中至少 3 个转录因子可同时作用于多个启动子,且兼具转录激活与抑制功能(图 2A)。 部分转录因子作用靶点高度专一:TcAP2-2 、TcMYB1 、TcNAC1 、TcNAC2 、bHLH2、TcWRKY1 分别仅靶向BAPT 、苯丙氨酸变位酶(PAM )、BAPT 、T5αH 、BAPT 与PAM 启动子。TcAP2-1 、TcAP2-3 、TcNAC4 、TcNAC5 对所有受试启动子均无调控作用(图 2A)。与之相反,有 9 个转录因子可结合多个启动子,具备广谱调控潜力。
研究还发现,TcMYB2 、bHLH1、TcNAC3 兼具转录激活与抑制双重功能,其作用效果取决于所结合的启动子类型。 在活性最强的前十组正向调控互作中,有 8 组由 MYB 家族蛋白介导,且多数靶向紫杉醇合成下游通路 基因的启动子。其中互作活性最强的组合为TcMYB3 与TASY ------TASY 编码的酶催化紫杉醇生物合成首个关键限速步骤 。
没有单一转录因子能够激活全部受试的紫杉醇合成基因启动子。但TcMYB3 可激活除T5αH 和BAPT 之外的所有启动子,而BAPT 被证实是紫杉醇合成通路的重要限速酶编码基因(5,6)。重要的是,TcMYB4 可特异性激活BAPT 启动子。
综上,TcMYB3 与 TcMYB4 协同作用 ,即可全面激活紫杉醇合成通路的核心基因启动子。
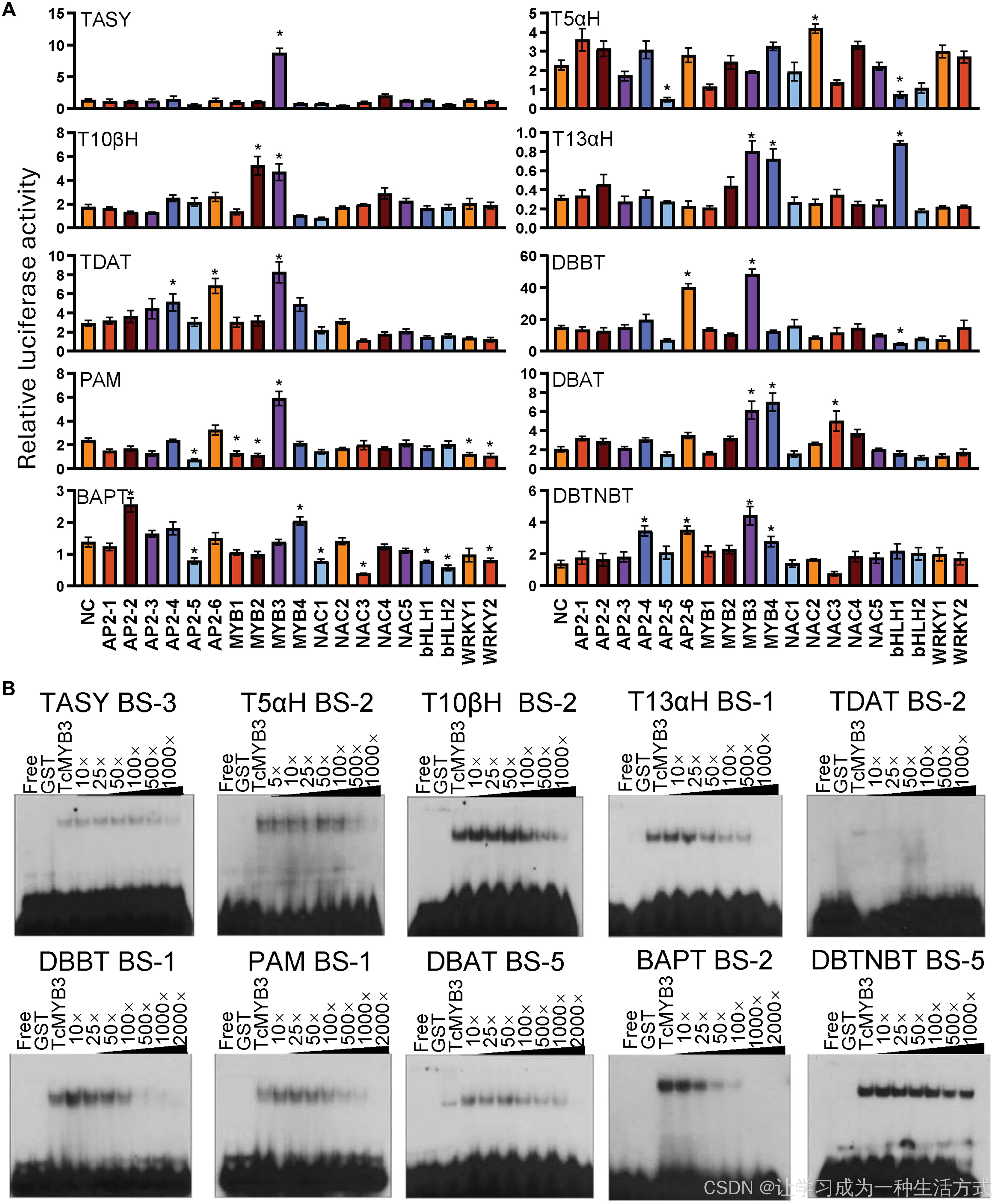
图 2 基于瞬时表达实验与凝胶迁移实验对候选转录因子的功能筛选
(A) 拟南芥瞬时表达实验(TEA):检测 19 个候选转录因子与 10 个紫杉醇生物合成基因启动子的互作关系。误差线代表标准误;生物学重复数 n ≥ 4;采用单因素方差分析结合图基检验,将各组与阴性对照组(NC)进行比较判定显著性,*P < 0.05。 (B) 竞争性凝胶迁移实验(EMSA):验证 TcMYB3 与 10 个紫杉醇合成基因启动子上结合位点(BS)的结合特异性。以谷胱甘肽 S - 转移酶(GST)作为阴性对照。各组 EMSA 电泳图上方标注了对应结合位点的寡核苷酸探针,探针详细序列见附表 S4。
凝胶迁移实验证实 TcMYB3、TcMYB4 可结合紫杉醇合成基因
瞬时表达实验提示 TcMYB3 与 TcMYB4 能够作用于紫杉醇合成基因启动子,为进一步验证二者可直接结合 靶标启动子,本研究表达并纯化获得两种转录因子的重组蛋白,开展凝胶迁移实验(EMSA)。
结果明确证实,TcMYB3 可强效结合全部 10 个紫杉醇合成关键基因的启动子序列,同时本研究在上述启动子中定位到 TcMYB3 的特异性结合位点(附图 S4)。竞争性凝胶迁移实验进一步验证了该结合作用的特异性(图 2B)。
TcMYB4 同样可稳定结合多个基因启动子,包括TASY 、DBAT 、DBBT 、DBTNBT 、BAPT ,以及编码关键早期酶(细胞色素 P450)的T5αH (16,17)。研究也在这些靶标启动子上鉴定出 TcMYB4 特异性结合位点,并通过竞争性 EMSA 验证了结合特异性(附图 S5)。附图 S6A 详细标注了两个转录因子在各启动子上的精确结合位置。
对结合序列进行分析发现,两类转录因子的结合区域均存在保守AC 元件 ,共有序列为 A (A/C) C(附图 S6B)。
综合实验结果:TcMYB3 可广泛结合 10 个核心合成基因的启动子,但在拟南芥瞬时表达体系中无法激活BAPT 与T5αH (图 2B);而 TcMYB4 可特异性结合这两个基因的启动子,恰好弥补了 TcMYB3 的调控短板。由此证实,TcMYB3-TcMYB4 蛋白组合 是协同调控紫杉醇生物合成通路的核心正向调控单元。
体内实验验证 TcMYB3、TcMYB4 可激活紫杉醇合成基因
为验证两个转录因子在红豆杉细胞内能否激活紫杉醇相关基因表达,本研究借助莫比乌斯组装技术 ,将报告基因与内参基因整合至同一载体,构建双荧光素酶报告载体(附图 S7A--C)。
首先优化红豆杉形成层分生细胞(CMC)的原生质体转染体系:结果显示,转染 DNA 用量为 16 μg 时瞬时表达效率最高;泛素 10 启动子(Ubiq10p)的转录活性显著强于 35S 启动子与 MAS 启动子(附图 S7D、E)。
基于优化后的红豆杉原生质体瞬时表达体系开展实验,结果与 EMSA 结论一致:TcMYB3 可激活全部 10 个紫杉醇合成关键基因的启动子(图 3A)。但该结果与异源拟南芥体系的实验结果存在差异 ------ 在拟南芥中,TcMYB3 无法激活BAPT 与T5αH (图 2A),这也反映出异源表达体系的局限性,推测原因是拟南芥细胞缺失红豆杉特有的辅助因子或互作蛋白。
此外,实验发现 TcMYB4 在红豆杉细胞中也可显著激活所有受试启动子(图 3A),该结果同样不同于其在拟南芥瞬时表达体系及 EMSA 中的表现。EMSA 证实 TcMYB4 能够结合TASY 与T5αH 启动子,但在拟南芥细胞中却无法激活这两个基因(图 2A、附图 S5),说明仅依靠 DNA 结合能力,不足以在异源拟南芥细胞中发挥转录激活功能。
以上结果表明,TcMYB3/TcMYB4 组合 的完整转录激活功能,高度依赖红豆杉自身的细胞调控体系,也凸显了在物种内源体系中开展转录因子功能研究的必要性。
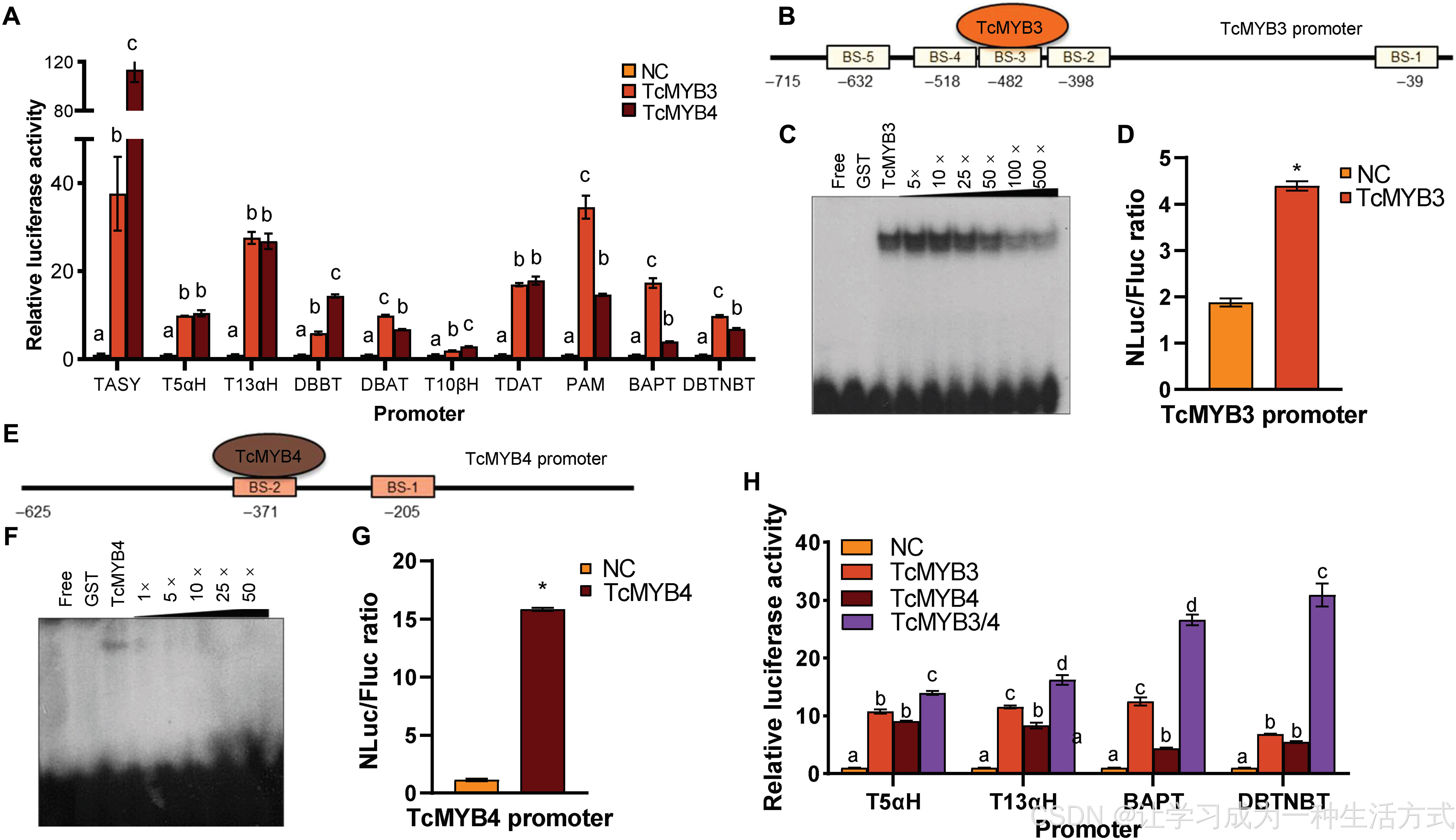
(A) 红豆杉形成层分生细胞原生质体瞬时表达实验:泛素 10(Ubqi10)启动子驱动的TcMYB3 、花椰菜花叶病毒 35S(CaMV 35S)启动子驱动的TcMYB4 ,分别对 10 个紫杉醇合成基因启动子的激活效果。 (B) TcMYB3 启动子结构示意图,标注其上 5 个 MYB 结合位点(MYB-BS)的位置。 (C) 竞争性凝胶迁移实验(EMSA),验证 TcMYB3 与自身启动子上 3 号结合位点(BS-3)的结合特异性;探针序列详见附表 S4。 (D) 红豆杉形成层分生细胞原生质体瞬时表达实验,验证全长TcMYB3 启动子(715 bp)介导的 TcMYB3 自激活效应。 (E) TcMYB4 启动子结构示意图,标注 MYB 结合位点位置。 (F) 竞争性凝胶迁移实验,验证 TcMYB4 与自身启动子上 2 号结合位点(BS-2)的结合特异性;探针序列详见附表 S4。 (G) 红豆杉形成层分生细胞原生质体瞬时表达实验,验证全长TcMYB4 启动子介导的 TcMYB4 自激活效应。 (H) TcMYB3 与 TcMYB4 共同转激活T5αH 、T13αH 、BAPT 、DBTNBT 启动子的实验结果。组间互作强度以NLuc/FLuc 比值 表示,均依据各组启动子对应的阴性对照组(NC)进行标准化;样本数 n > 3。(D)(G) 组采用独立样本t 检验,与阴性对照相比,*P < 0.05。数据以平均值 ± 标准差 呈现,n =3;不同字母代表组间存在显著差异(P < 0.05)。
上述实验结果存在明显差异,因此本研究进一步探究TcMYB3、TcMYB4 的自调控作用 以及二者之间的相互转激活关系。 以欧洲红豆杉形成层分生细胞为材料开展瞬时表达实验,结果显示:TcMYB3 可激活自身截短至−398 bp、−525 bp 的启动子片段,但无法激活−258 bp 的截短启动子(图 3B、附图 S8A、S8B)。TcMYB3 启动子−525 bp 至−258 bp 区间内,在−518 bp、−482 bp、−298 bp 位置预测到 3 个 MYB 结合位点(图 3B)。凝胶迁移实验证实,TcMYB3 可特异性结合−482 bp 处的序列AACACC(图 3B、3C,附图 S8A、S8B)。欧洲红豆杉细胞瞬时表达实验也验证了全长TcMYB3 启动子可介导该基因自激活(图 3D)。
研究同时在TcMYB4 启动子上检测到自激活现象:全长−625 bp 启动子具备自激活活性,截短至−278 bp 后该活性消失(图 3E、附图 S8C)。凝胶迁移实验证明,TcMYB4 的结合区域位于其启动子−371 bp 范围内(图 3E、3F,附图 S8D),全长启动子的自激活效应也经细胞瞬时表达实验得到验证(图 3G)。
此外,双因子互作实验表明:TcMYB4 可转激活TcMYB3 启动子片段,但 TcMYB3 无法反向调控TcMYB4 (附图 S9),说明二者存在层级调控关系 。
为明确两个 MYB 转录因子在激活紫杉醇合成酶基因启动子过程中的协同效应,本研究在欧洲红豆杉形成层分生细胞原生质体中开展共表达实验。结果显示,TcMYB3 与TcMYB4 共表达时,可显著增强对多个紫杉醇合成相关启动子的激活作用,其中以T5αH 、T13αH 、BAPT 、DBTNBT 这 4 个关键基因启动子的激活提升效果最为明显(图 3H)。以上结果证实,TcMYB3 与 TcMYB4 共转染可协同激活上述 4 个基因的表达 。
构建稳定过表达 MYB 转录因子的红豆杉细胞株
为验证TcMYB3 与TcMYB4 共表达能否在红豆杉细胞中持续驱动紫杉醇生物合成,本研究构建稳定转基因红豆杉形成层分生细胞株 。实验分别采用β- 雌二醇诱导型启动子 、地塞米松(Dex)诱导型启动子 构建TcMYB3 、TcMYB4 表达载体(附图 S10A)。经诱导处理后,可分别检测到两个基因的特异性表达(图 4A、附图 S10B--D)。
单独诱导表达TcMYB3 或TcMYB4 ,均可上调紫杉醇合成酶基因的转录水平(图 4B、附图 S11A),与原生质体瞬时表达结果一致(图 3A)。相较于单基因表达,两个基因共表达 可进一步显著提升合成酶基因的表达量,直接证明二者存在协同调控功能(图 4B)。
在转基因细胞株中,TcMYB3 与TcMYB4 共表达可大幅提升两种关键产物积累:商业化前体物质巴卡亭 Ⅲ(图 4C)与紫杉醇(图 4D)含量显著上升。其中,紫杉醇含量达 98 μg/g 鲜重,较对照组提升 121 倍;巴卡亭 Ⅲ 含量达 877 μg/g 鲜重,较对照组提升 364 倍。
对双基因过表达细胞株施加茉莉酸甲酯诱导后,紫杉醇与巴卡亭 Ⅲ 产量进一步增加(附图 S11B),体现出多重调控叠加效应。推测该现象一方面是茉莉酸甲酯激活了细胞内源 MYB3/4,另一方面可能还存在其他未鉴定的茉莉酸甲酯响应型激活因子参与调控。
综上,TcMYB3 与TcMYB4 共表达可有效促进红豆杉形成层分生细胞合成紫杉醇及其他紫杉烷类物质。同时本研究表明,茉莉酸甲酯介导的转录调控网络十分复杂,该通路可作用于 TcMYB3/TcMYB4 调控模块的上游、下游,或与之并行,共同调控紫杉烷类化合物的生物合成。
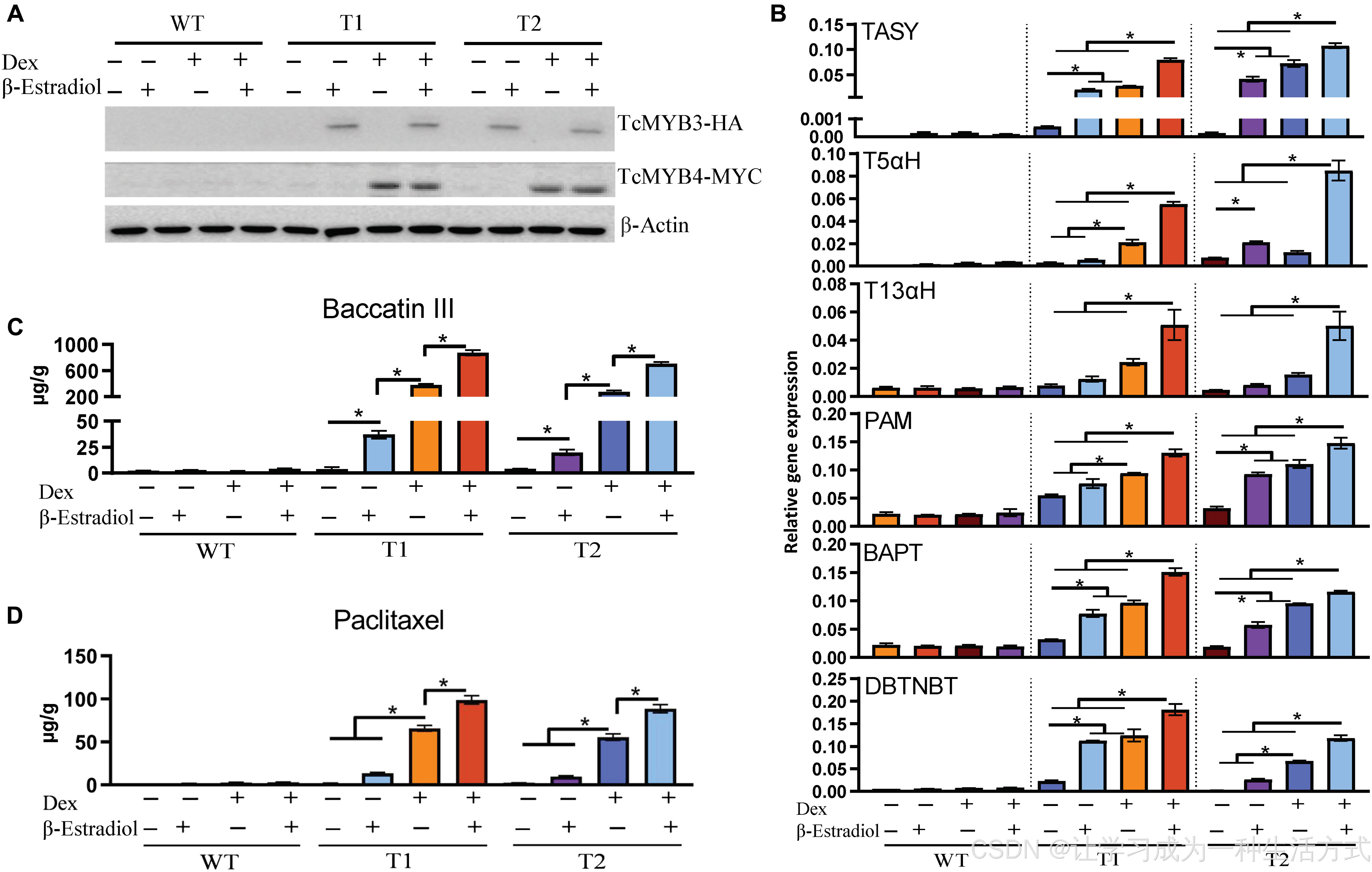
+(A) 采用血凝素(HA)抗体、MYC 抗体,分别检测转基因细胞株中 TcMYB3 与 TcMYB4 的蛋白表达水平;以 β- 肌动蛋白作为上样对照。样品经地塞米松、β- 雌二醇诱导后收集。WT:野生型细胞;T1、T2:转基因株系 1、2。 (B) 经对应化学物质处理 12 小时后,各株系中紫杉醇生物合成通路关键基因的转录表达水平。 (C、D) 经对应化学物质处理 7 天后,细胞中巴卡亭 Ⅲ(C)与紫杉醇(D)的产量,单位为微克 / 克细胞鲜重。数据以平均值 ± 标准差 表示,包含 3 组生物学重复;*P < 0.05。
讨论
目前商业化紫杉醇生产依赖半合成工艺,该方法成本高昂且环境友好性差,需要从红豆杉植株中提取巴卡亭 Ⅲ 等关键中间产物。茉莉酸甲酯是经典的植物天然产物诱导子,现已应用于工业化植物细胞培养体系,用以提升紫杉醇产量 (20,25)。挖掘茉莉酸甲酯诱导下、可上调紫杉醇合成的调控因子,能够为细胞培养体系的遗传改造、提高紫杉醇产能提供理论依据。
本研究解析了茉莉酸甲酯促进红豆杉形成层分生细胞合成紫杉醇的转录调控机制,鉴定出TcMYB3-TcMYB4 调控模块 。该模块可作为核心调控开关,同步激活紫杉醇合成通路中 10 个关键基因的启动子(图 2、附图 S1E)。在稳定表达该模块的转基因红豆杉细胞中,紫杉醇产量可达 98 微克 / 克鲜重(图 4D),与茉莉酸甲酯诱导组产量(102 微克 / 克鲜重)基本持平 (20)。这一结果标志着相关研究从解析复杂细胞应答,迈向精准转录调控系统的人工设计 。
本团队前期研究证实,红豆杉形成层分生细胞在规模化培养过程中代谢稳定性优异:摇瓶培养经诱导 45 天后,紫杉醇产量为 102 毫克 / 千克;20 升气升式生物反应器培养体系中,产量可提升至 268 毫克 / 千克 (20)。国外药企菲顿生物科技目前工业化体系的紫杉醇产量约为 1~2 克 / 千克,但这类专利体系依托高度优化的大型生物反应器,且诱导方案均针对专属细胞株精细调试。
学界已对短叶红豆杉、欧洲红豆杉、东北红豆杉、中国红豆杉及曼地亚红豆杉(欧洲红豆杉与东北红豆杉杂交种)等多个物种开展紫杉醇及紫杉烷类物质合成研究 (25),工业化生产多选用中国红豆杉或曼地亚红豆杉的高产细胞株。本研究采用欧洲红豆杉构建 MYB 过表达细胞株。需要说明的是,受细胞筛选历史、工艺优化程度影响,不同物种、不同细胞株的产能差异极大 。工业产能数据,是大型生物反应器经过完整生产周期(通常 3~5 周)优化后得到的最终产量;而本研究中转基因细胞的产量,仅来自小规模、未经过工艺优化的摇瓶培养,且仅检测了诱导 7 天这单一时间点(图 4D)。因此,本研究现阶段实验室体系的产量,无法直接与工业化细胞工厂的产能对标。
本研究的核心价值在于:通过基因改造实现产物产量较对照组大幅提升 ,为后续工艺优化奠定了基础。该研究发掘的 TcMYB3/4 遗传工具,可应用于各类高产细胞株以提升产能,不受物种来源与前期优化背景的限制。
相比传统提取工艺,红豆杉形成层分生细胞培养体系原料供给可持续、生物量近乎无限。本研究鉴定的 R2R3-MYB 转录因子,填补了当前紫杉醇产量提升的技术短板。通过诱导型系统过表达这类正向调控因子,可在细胞株长期传代过程中稳定维持高产表型 ,保障培养体系与细胞活性。本研究采用 β- 雌二醇诱导系统完成概念验证,若面向临床级工业化生产,建议替换为地塞米松或乙醇诱导系统,可规避雌激素受体阳性相关医疗场景带来的监管风险 (26)。
已有研究在异源宿主中实现了紫杉醇及其工业前体巴卡亭 Ⅲ 的从头生物合成 (12)。目前,巴卡亭 Ⅲ 与紫杉醇合成所需的全部关键酶(包括此前多个 "缺失基因")均已被鉴定,异源宿主中完整的巴卡亭 Ⅲ 合成通路也已成功重构 (5,15,27)。例如,研究者在本氏烟草中表达最简合成基因组合,巴卡亭 Ⅲ 产量可达 154.84 纳克 / 克,接近红豆杉针叶中的天然含量(225.32 纳克 / 克)(18);另有团队在烟草中导入 17 个合成相关基因,巴卡亭 Ⅲ 积累量同样与红豆杉针叶天然水平相当 (5)。
尽管紫杉醇合成通路已在异源宿主中完成解析与重构,但该技术落地应用仍存在瓶颈:异源体系难以模拟植物内源的代谢流调控,且细胞色素 P450 酶普遍存在表达效率低的问题 (12,17,28,29)。本研究结果也印证了这一点:TcMYB3 在红豆杉内源体系中可激活所有关键基因启动子,但在拟南芥瞬时表达体系中,却无法激活BAPT 与T5αH 启动子(图 2A)。推测原因是拟南芥缺失红豆杉特有的辅助因子与互作蛋白,体现了异源表达系统的局限性。
相较之下,红豆杉原生细胞培养仍是极具潜力的发展方向,兼具可持续性与环境友好性,也便于开展代谢工程改造以实现规模化生产 (19)。已有研究表明,培养条件优化结合诱导处理,可使紫杉醇产量达到 43.43 毫克 / 升 (30);在欧洲红豆杉细胞中过表达BAPT 与DBTNBT 基因,紫杉烷类总产量可达 53.83 毫克 / 升 (31)。本研究鉴定出紫杉醇合成的核心转录因子网络,改造后的转基因红豆杉细胞紫杉醇产量达 98 微克 / 克鲜重,较对照组提升 121 倍;关键前体巴卡亭 Ⅲ 产量达 877 微克 / 克鲜重,增幅达 364 倍(图 4C、4D),产量显著优于现有异源表达体系与其他细胞培养体系的报道结果 (32)。
转录因子改造是提升植物代谢产物产量的高效策略。该技术已成功应用于多种场景:调控真菌聚酮化合物通路以生产洛伐他汀、重构花青素通路培育高花青素番茄等 (33-35)。以往针对紫杉醇合成的基因改造,多聚焦于单一转录因子过表达,例如 MYB、bHLH、NAC 家族蛋白,包括近年报道的中国红豆杉 MYB29a、曼地亚红豆杉 TmMYB3、TmMYB39-TmbHLH13 等 (32,36,37)。这类研究虽明确了部分调控靶点,但难以实现多酶协同表达,无法支撑产物高水平合成。究其原因,单一基因改造无法在异源体系中同步激活整条合成通路 (13)。
本研究突破了传统思路,无偏筛选并构建了适配合成通路的转录因子网络 。这种组学层面的筛选策略,摆脱了单基因靶向研究的局限,筛选出一组最简转录因子组合,可协同激活整条代谢通路。细胞内功能验证证实,该转录因子网络可显著提升紫杉醇合成能力(图 2、3),具备很高的实际应用价值与发展潜力。
本研究的核心创新在于:不再局限于已知转录因子的功能解析,而是从头筛选并构建适配产物合成的转录调控网络。这种多因子协同调控模式,比传统单酶改造策略调控效率更高、协同性更强,也标志着植物代谢调控研究正式迈入系统调控 新阶段。
本研究虽已大幅提升紫杉醇及巴卡亭 Ⅲ 产量,但基于现有成果,仍有多条技术路线可进一步优化产能。TcMYB3-TcMYB4 模块足以强效诱导红豆杉细胞合成紫杉醇与紫杉烷类物质,但该模块并非茉莉酸甲酯诱导通路中唯一的核心调控因子。实验发现,在 TcMYB3/4 过表达株系中施加茉莉酸甲酯,紫杉醇与巴卡亭 Ⅲ 积累量会进一步提升(附图 S11B),说明细胞内存在更复杂的转录调控网络:还有大量未鉴定的茉莉酸甲酯响应型转录因子,可作用于 TcMYB3/4 模块的上游、下游或与之平行,共同完成通路的全激活。
茉莉酸信号通路核心调控因子 bHLH 类转录因子 AtMYC2 (38),是后续机制研究的重点候选协同因子。结合本研究成果,后续可从以下方向进一步提升产能: 第一,深入解析 TcMYB3-TcMYB4 调控网络,鉴定其互作蛋白,明确茉莉酸甲酯诱导下转录因子的结合调控机制,挖掘可协同或抑制通路活性的内源调控因子,为后续基因改造提供靶点; 第二,鉴定通路中的内源抑制因子,利用 CRISPR-Cas9 基因敲除技术解除抑制作用,强化合成通路活性;同时敲除苯丙烷通路等竞争支路,引导代谢流定向流向紫杉醇合成通路 (39,40); 第三,将现有遗传改造方案与培养体系优化相结合,推动技术规模化落地。
综上,本研究证实转录因子网络定向改造 是一套成熟可行的核心策略,也为复杂植物天然产物的可持续生产开辟了全新方向。
总而言之,本研究验证了转录网络改造在提升高价值植物次生代谢产物产量中的应用潜力。我们认为,通过系统性筛选与解析其他转录因子网络,有望为各类天然产物打造稳定、高产的规模化生产体系。
材料与方法
转录组分析
采用 DESeq2 对含生物学重复的数据集进行差异表达分析 (41)。校正后P 值(Padj)<0.05、对数二倍表达变化绝对值(|log2FC|)>1 的基因,判定为茉莉酸甲酯处理组的差异表达基因(DEG)。以全部注释转录本为背景进行功能富集分析,采用费希尔精确检验 结合本雅明--霍赫贝格错误发现率(FDR) 校正评估显著性。利用VennDiagram包绘制韦恩图,展示不同时间点差异基因的交集;以柱状图统计各时间点差异基因数量。
整合所有差异基因,基于 Z 分数标准化的 log2FC 值,借助ComplexHeatmap包绘制热图,并采用欧氏距离进行聚类。采用K 均值聚类( k =6) 对差异基因进行时序表达模式划分,通过平滑折线图展示各类群基因的 Z 分数表达趋势。分别针对生物学过程(BP)、分子功能(MF)、细胞组分(CC) 开展富集分析,统计方法同上,并将排名前十的显著 GO 条目绘制成气泡图。
基因组 DNA 提取
取约 100 mg 冷冻样品,液氮充分研磨后加入提取缓冲液,65℃水浴 30 min。加入等体积氯仿--异戊醇混合液(体积比 24:1),混匀后 4℃、10000g 离心 10 min。吸取上清并重复抽提,加入 1.5 倍体积异丙醇,−20℃静置 1 h 沉淀 DNA。4℃、17000g 离心 30 min 收集 DNA 沉淀,70% 乙醇洗涤,室温风干后加入 50 μL Tris-EDTA(TE)缓冲液,65℃孵育 5 min 充分溶解。
cDNA 末端快速扩增
采用cDNA 末端快速扩增技术(RACE) 扩增编码区不完整基因的全长序列。对TcAP2-3 、TcNAC3 、TcbHLH2 进行 5' RACE 扩增,对TcMYB4 进行 3' RACE 扩增。使用罗氏第二代 5'/3' RACE 试剂盒,按照说明书完成目的片段扩增、克隆及测序,引物序列见附表 S1。
基因表达分析
按照图注时间点收集东北红豆杉形成层分生细胞(CMC),采用改良十六烷基三甲基溴化铵(CTAB)法 提取总 RNA (42)。总 RNA 经脱氧核糖核酸酶处理后,以 oligo (dT) 为引物,使用赛默飞第一链 cDNA 合成试剂盒反转录得到 cDNA。采用罗氏 LightCycler 480 实时荧光定量 PCR 仪进行 qRT-PCR 检测,所用基因特异性引物见附表 S2 及参考文献 (43)。每组实验设置 3 次技术重复与 3 组生物学重复。以红豆杉肌动蛋白基因作为内参基因校正相对表达量,采用单因素方差分析结合图基检验进行显著性分析(P <0.05)。
拟南芥瞬时表达载体构建
将 pRL-TK 载体(普洛麦格)的海肾荧光素酶基因片段插入 p2GW7.0 载体,构建内参对照载体。使用高保真 DNA 聚合酶(纽英伦生物技术),以 cDNA 为模板扩增转录因子编码序列,以基因组 DNA 为模板扩增基因启动子片段。将转录因子序列插入 p2GW7.0 构建效应载体 ;将启动子片段插入 pGWlucB 载体的萤火虫荧光素酶基因上游,构建报告载体 。引物序列见附表 S3。
拟南芥原生质体瞬时表达实验
利用 Gateway 克隆系统完成 10 个红豆杉启动子与 19 个转录因子的载体构建(附表 S3)。原生质体转染参照已优化的实验方案 (44)。取 250 μL 原生质体悬液(浓度 5×10⁵个 /mL)与 14 μL 质粒混合液(浓度 1 μg/μL;效应载体:报告载体:对照载体体积比 = 4:5:5),加入聚乙二醇--钙离子溶液(含 40% PEG 4000、0.4 M 甘露醇、0.1 M 硝酸钙,pH 8.0~9.0),室温孵育 5 min。
洗涤细胞后,置于改良 W5 溶液中,25℃光照条件下培养 20 h;液氮速冻后−80℃保存。采用普洛麦格双荧光素酶检测系统测定酶活,实验重复 3 次。与空载体对照组相比酶活显著升高(P <0.05),判定为存在正向互作。
重组 TcMYB3、TcMYB4 蛋白的表达与纯化
依据附表 S3 引物,将TcMYB3 、TcMYB4 编码序列克隆至带谷胱甘肽 S - 转移酶(GST) 标签的 pDEST24 载体,转化大肠杆菌 BL21 (DE3) 菌株。当菌液 OD₆₀₀达到 0.6~0.7 时,加入 0.1 mM 异丙基 -β-D - 硫代半乳糖苷,28℃诱导蛋白表达 3 h。
菌体裂解液配方:25 mM Tris-HCl(pH 7.5)、100 mM 氯化钠、0.1% Triton X-100、1 mg/mL 溶菌酶、1 mM 二硫苏糖醇(DTT)及蛋白酶抑制剂。使用 GE Healthcare 谷胱甘肽琼脂糖 4B 树脂纯化 GST 融合蛋白,单独表达的 GST 蛋白作为阴性对照。
电泳迁移率变动分析(EMSA)
选取紫杉醇合成基因启动子中含顺式作用元件的序列,合成长度 20 bp 的双链寡核苷酸探针(附表 S4)。利用 T4 多核苷酸激酶,以 γ-³²P ATP 对探针末端进行放射性标记。
结合反应体系:0.5 μg 重组蛋白、20 mM HEPES(pH 7.9)、50 mM 氯化钾、5 mM 氯化镁、1 mM DTT、5% 甘油、0.1 μg/μL 聚脱氧肌苷 - 脱氧胞苷、50 μg/μL 牛血清白蛋白(BSA)。4℃孵育 20 min,室温继续反应 20 min。
反应产物上样至 6% 非变性聚丙烯酰胺凝胶,1×Tris - 硼酸 - EDTA 缓冲体系、120 V 电泳 80 min。凝胶烘干后,−80℃条件下与 X 光胶片过夜曝光显影。
热不对称交错 PCR(TAIL-PCR)
参照 Thanh 等人的方法 (45),采用热不对称交错 PCR 扩增TcMYB3 与TcMYB4 的启动子序列。取 50 ng 东北红豆杉 CMC 基因组 DNA,利用嵌套基因特异性引物(RSP1、RSP2、RSP3)与简并随机引物(附表 S5)进行三轮连续扩增。扩增产物连接至 pGEM-T Easy 载体(普洛麦格)并测序。将获得的启动子序列再次扩增,插入莫比乌斯克隆通用受体载体。
莫比乌斯载体组装
采用莫比乌斯组装技术 (24) 构建载体,分别用于红豆杉原生质体瞬时表达与细胞稳定转化。将启动子与纳米荧光素酶(NLuc)融合;TcMYB3 、TcMYB4 分别由 PLexA、Ppop6 启动子驱动。将 3 个一级转录表达单元组装为二级载体,载体包含转录因子(TcMYB3/TcMYB4)、启动子 - NLuc 融合片段及萤火虫荧光素酶(FLuc)报告基因(附图 S7)。稳定转化所用载体见附图 S10A。
红豆杉 CMC 原生质体制备与转染
取 1 mL 压实的东北红豆杉 CMC 细胞,加入 10 mL 酶解液(MGG 缓冲液配制,含 1% 纤维素酶 R-10、0.25% 果胶酶 Y-23、0.05% BSA、0.01% MES),避光酶解 3 h。酶解产物经 40 μm 尼龙滤网过滤,100 g 离心 3 min 收集原生质体,预冷 MGG 溶液洗涤 2 次。
冰上重悬于 1 mL MGG 溶液静置 1 h,调整浓度至 5×10⁵个 /mL(MMM 溶液:0.5 M 甘露醇、15 mM 氯化镁、0.1% MES)。采用伊文思蓝与荧光素二乙酸染色检测细胞活性。
取 250 μL 原生质体悬液、8 μg 质粒与 250 μL PEG 溶液(40% PEG 4000、0.4 M 甘露醇、0.1 M 硝酸钙)混合,室温转染 1 min;加入 MGG 溶液稀释,避光静置 1 h。洗涤后重悬于 500 μL MGG 溶液,过夜培养后检测荧光素酶活性。
双荧光素酶活性检测
使用普洛麦格 Nano-Glo 双荧光素酶检测试剂盒测定荧光素酶活性。向样品中加入 80 μL 细胞裂解液,依次添加 80 μL 萤火虫荧光素酶检测试剂、80 μL 纳米荧光素酶终止检测试剂。采用伯腾多功能酶标仪检测发光值(增益 3600,积分时间 1 s),以 NLuc/FLuc 比值并结合对照组进行数据标准化。实验重复 2 次,每组设置 3 个生物学重复,采用t 检验分析显著性(P <0.05)。
红豆杉 CMC 稳定转化与蛋白免疫印迹
农杆菌 AGL1 菌株接种于 YEP 液体培养基(含 25 mg/L 利福平、50 mg/L 卡那霉素),28℃培养 2~3 d。取 1 mL 菌液转接至 20 mL 含对应抗生素与 200 μM 乙酰丁香酮的 YEP 培养基,培养至 OD₆₀₀=0.8。3270 g 离心 5 min 收集菌体,重悬于侵染缓冲液(10 mM MES、10 mM 氯化镁、250 μM 乙酰丁香酮,pH 5.6),22℃静置 1~4 h。
加入 0.02% Silwet-77 表面活性剂,取 50 mL 菌液与 50 mL 红豆杉 CMC 细胞混合,25℃、110 rpm 避光振荡 30 min。无菌水洗涤 2 次,转移至 TB 培养基共培养 3 d。多次洗涤后涂布于含 400 μM 替考拉宁的 TB 固体培养基,培养 7 d;随后逐步降低替考拉宁浓度(300~100 μM)、提升潮霉素浓度(10~25 μg/L)进行梯度筛选,获得稳定转化细胞株。
采用免疫印迹法检测目的蛋白:一抗为 Myc 标签抗体(稀释比 1:2000)、HA 标签抗体(稀释比 1:2000);二抗为 IRDye 800CW 标记羊抗鼠 / 羊抗兔抗体(稀释比 1:5000)。
紫杉烷类物质提取与定量检测
按照图注时间,采用 50 μM β- 雌二醇、50 μM 地塞米松或 100 μM 茉莉酸甲酯处理 CMC 细胞。取约 50 mg 冷冻细胞,加入 1 mL 甲醇--二氯甲烷混合液(体积比 1:1),超声提取 2 h。真空浓缩上清液,残留物用二氯甲烷--水(体积比 2:1)重复萃取 2 次。合并有机相并干燥,甲醇复溶后进行液相色谱--质谱联用(HPLC-MS)检测。
采用安捷伦 1290 超高效液相色谱串联 6560 离子淌度四极杆飞行时间质谱仪,搭配 ACE C18-PFP 色谱柱(100×2.1 mm,粒径 1.8 μm)进行定量分析。流动相 A 为纯水、B 为甲醇,均添加 0.1% 甲酸;洗脱梯度:7 min 内 B 相比例由 10% 升至 100%,再等度洗脱 4 min,流速 0.3 mL/min,柱温 30℃。
质谱采用正离子模式,质荷比扫描范围 400~1000,采集频率 1 帧 / 秒。监测准分子离子峰:巴卡亭 Ⅲ M+Na⁺ = 609.2306,紫杉醇 M+Na⁺ = 876.3202。采用标准品(剑桥生物科学公司)绘制标准曲线,完成样品定量。