
摘要
温和噬菌体(Temperate bacteriophages)是与细菌宿主共同进化的普遍存在的病毒,其定义特征在于能够经历两种截然不同的生命周期:裂解循环(lytic cycle)和溶原循环(lysogenic cycle)。长期以来,温和噬菌体一直是微生物学、遗传学和进化生物学研究的基础模型,其生命周期是病毒学中表征最为透彻的领域之一。历史上,噬菌体生命周期主要从基因组切除、复制和包装如何驱动感染性颗粒形成的角度来审视。尽管这一视角抓住了噬菌体生命周期的核心过程,但这种狭隘的观点忽略了噬菌体与宿主以及其他可移动遗传元件(mobile genetic elements, MGEs)之间全方位的相互作用。在这篇综述中,作者基于新出现的、扩展了该框架并带来意外复杂性的见解,重新审视了温和噬菌体的生命周期。作者认为,噬菌体(原噬菌体)的许多特性应被视为其生命周期的组成部分,而非单独讨论。这种整体性观点对于充分理解温和噬菌体生命周期的复杂性以及这些病毒在微生物学和生物技术应用中的关键作用至关重要。
- Rostøl, J. T., Chmielowska, C., Marina, A., & Penadés, J. R. (2026). Revisiting the life cycle of temperate phages. Nature Reviews Microbiology. https://doi.org/10.1038/s41579-026-01318-7
- 期刊:Nature Reviews Microbiology (IF=103.3)
- 发表时间:2026年5月20日
引言
噬菌体(Bacteriophages, phages)是依赖细菌宿主完成其生命周期的病毒。在一个多世纪前被发现后,噬菌体如今已知是地球上最丰富的生物实体,估计有10^31个颗粒,数量超过细菌10倍。噬菌体是细菌生存、致病性和进化的关键参与者。历史上,它们一直是分子生物学研究的主力军,在诸如随机突变如何导致进化适应以及核酸是遗传物质等基础发现中处于核心地位。此外,噬菌体研究还产生了多种生物技术工具,如限制性内切酶、CRISPR-Cas编辑系统以及用于mRNA疫苗生产的T7 RNA聚合酶。
大多数噬菌体由封装在蛋白质结构中的核酸遗传物质组成。噬菌体核酸必须被递送到细菌内部,在那里它可以劫持宿主机制进行噬菌体繁殖。然而,除了这些基本共同的结构和过程外,噬菌体在形态、遗传组成、遗传组织和生命周期方面可能高度多样化。因此,除了作为专性细菌寄生虫外,很难确定噬菌体之间共有的许多特性,这使得有必要研究不同的噬菌体模型以深入了解其全部多样性。
双链DNA温和噬菌体,特别是原型大肠杆菌噬菌体λ,历来备受关注。温和噬菌体遵循两种截然不同的生命周期之一。在裂解循环中,噬菌体感染宿主以产生更多病毒拷贝,确保其水平传播。在溶原循环中,噬菌体进入宿主细胞内一种更为被动的状态,通常(但并非总是)通过将噬菌体基因组整合到细菌染色体中实现。在此,噬菌体作为原噬菌体(prophage)随宿主DNA被动复制,确保其垂直传播,直到被重新激活(诱导)再次进入裂解循环。从词源学上讲,"溶原菌"(lysogen)是指能够引起其他菌株裂解(因为释放噬菌体)的细菌菌株,即携带温和原噬菌体的菌株。噬菌体λ在裂解和溶原化之间的决策,通常被称为"遗传开关"(genetic switch),几十年来一直是研究遗传电路调控的模型。人们曾假设其他温和噬菌体的生命周期遵循类似的通用模式。
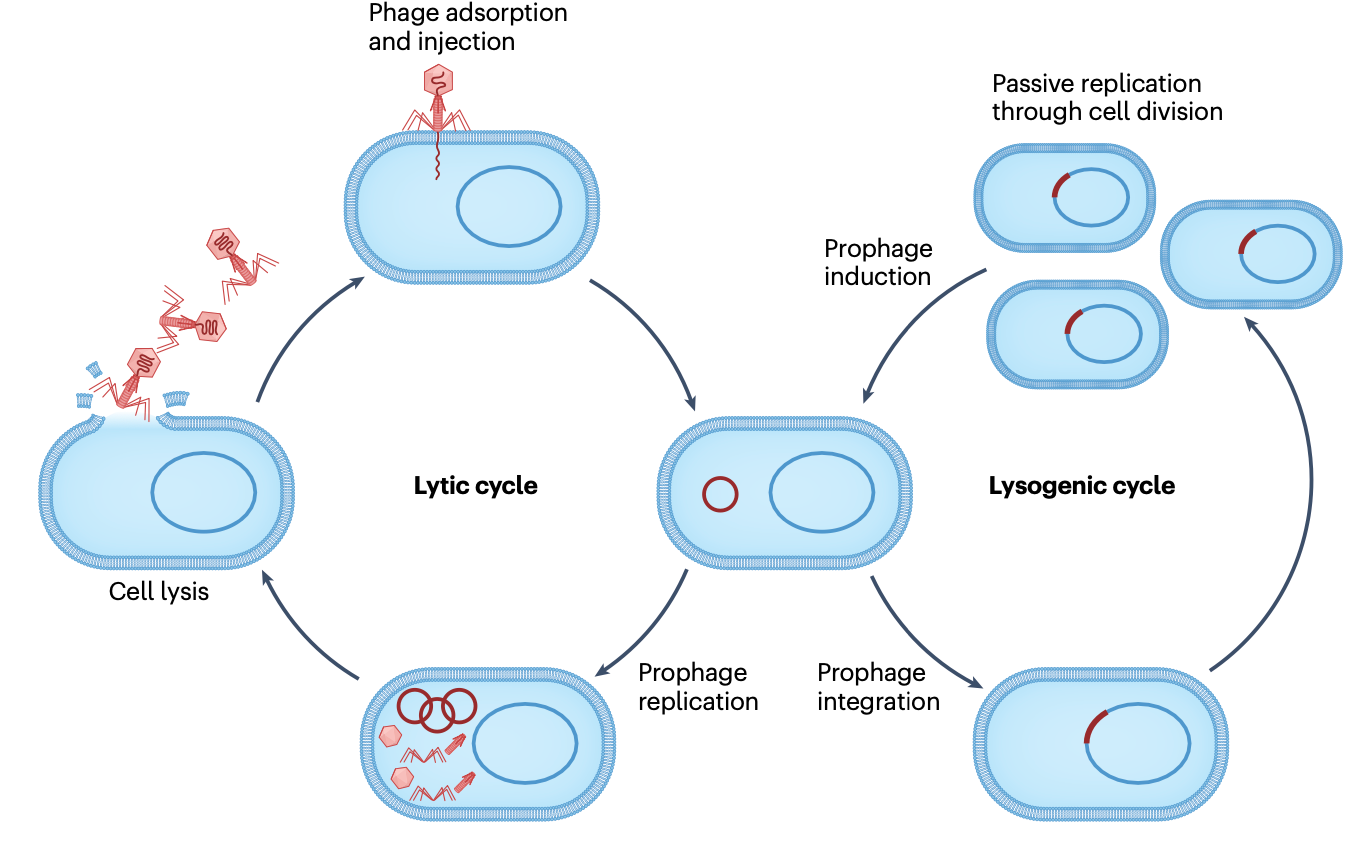
历史上,大多数研究集中在三个对产生感染性颗粒至关重要的核心过程上,即噬菌体基因组切除与整合、复制和包装,以及这些核心过程如何被调控。然而,研究进展使这种对温和噬菌体生命周期的相当直接的看法变得复杂,揭示了新的机制和已知过程的新方面,这些方面深刻地塑造了噬菌体生物学、繁殖及其与宿主的相互作用。
在这篇综述中,作者描述了对温和噬菌体生命周期的更新观点。作者与他人共同提出,温和噬菌体生命周期的定义应比以往公认的更广泛。尽管历史上的研究重点主要集中在一小部分噬菌体的分子机制上,但当代观点应认识到分子机制与更广泛多样噬菌体的生态和进化之间的联系。在这种当代观点中,许多间接因素决定了温和噬菌体的生存、适应性和繁殖,例如原噬菌体基因在溶原期间对宿主表型的贡献、影响裂解与溶原化决策的群体感应、替代诱导信号,以及感染期间对细胞防御的规避。此外,原噬菌体诱导过程中事件的时间顺序可能与以往认为的不同,这对细菌群落具有重要影响。最后,作者讨论了温和噬菌体生命周期与其他可移动遗传元件生命周期之间相互作用的重要性。
原噬菌体阶段(非休眠状态)
温和噬菌体的一个标志是它们可以作为原噬菌体存在于细胞内,并与宿主细菌一起被动复制。从噬菌体的角度来看,溶原化是有益的,因为原噬菌体DNA复制不受细菌免疫系统的阻碍。此外,溶原化意味着噬菌体不需要反复寻找新的、可能稀少的宿主来支持裂解生命周期。溶原化也为宿主细菌提供了优势,因为原噬菌体可以通过多种方式调节宿主适应性和行为。这些优势包括提供对外部噬菌体捕食的抗性、增强毒力和致病性,以及影响各种宿主过程,如生物膜形成、孢子形成、运动性和群体感应。这种互利共生宿主-噬菌体关系背后的逻辑是,通过增强宿主的适应性,原噬菌体也增加了其自身在环境中生存和传播的概率。
大多数已测序的细菌基因组至少含有一个原噬菌体,并且细菌基因组的相当大比例可以由原噬菌体DNA组成。越来越清楚的是,原噬菌体在溶原化过程中扮演着活跃的角色,而不仅仅是依附于其细菌宿主的寄生虫。这种对宿主适应性的贡献在许多情况下可以被认为是原噬菌体生存的内在部分,因此是温和噬菌体生命周期的组成部分。
原噬菌体通过在溶原循环期间表达特定基因来促进宿主生理。历史上,此类基因的研究与其在细菌毒力和发病机制中的作用相关。原噬菌体含量的增加通常会导致毒力增强,例如在慢性肺部感染的大鼠模型中,缺失各种原噬菌体元件的利物浦流行株铜绿假单胞菌突变体的竞争力下降所证明。有助于细菌生理学的基因包括"噬菌体冗余基因"(phage morons,即拥有自身启动子、为噬菌体基因组"增添更多"的基因)、从溶原启动子表达的基因以及编码非编码RNA的基因。由于许多温和噬菌体能够轻易在细菌菌株之间转移,细菌可以通过获得带有不同附属基因的新原噬菌体来快速适应新的生态位。
原噬菌体可以通过携带编码增强细菌毒力的毒素基因来增加宿主适应性,包括白喉棒状杆菌(Corynebacterium diphtheriae)中的白喉毒素和金黄色葡萄球菌(Staphylococcus aureus)中的Panton-Valentine杀白细胞素。原噬菌体还可以在感染期间帮助其致病宿主生存。例如,噬菌体starP-Sa3int编码酶TarP,该酶修饰金黄色葡萄球菌表面的壁磷壁酸,促进免疫逃逸。类似地,通过表达超氧化物歧化酶SodC,沙门氏菌噬菌体Gifsy-1保护细菌在巨噬细胞内免受超氧化物介导的杀伤。
重要的是,许多原噬菌体附属基因编码抗噬菌体防御系统,保护溶原菌免受外部噬菌体捕食。因此,原噬菌体被认为是宿主细菌抗噬菌体库的主要贡献者,并促进防御系统在整个细菌种群中的传播。对大肠杆菌基因组中抗噬菌体系统位置的图谱绘制显示,大多数热点区域(即抗噬菌体系统富集的基因组位置)是整合的原噬菌体。研究表明,在许多情况下,单个原噬菌体通过多种机制提供对多种噬菌体感染的强大抗性,例如在铜绿假单胞菌、耻垢分枝杆菌(Mycobacterium smegmatis)、大肠杆菌和戈登氏菌属(Gordonia spp.)中。这些发现突显了防御基因在原噬菌体中的高发生率。例如,大肠杆菌HK97噬菌体的gp15通过阻断噬菌体DNA注入来保护细菌免受类似噬菌体的感染。
噬菌体还可以通过"活性溶原"(active lysogeny)调节宿主致病性,在此过程中,它们通过整合到染色体基因中或从其中切除来充当调控开关。例如,单核细胞增生李斯特菌(Listeria monocytogenes)中的comK基因通常被整合的原噬菌体φ10403S中断。当致病性单核细胞增生李斯特菌感染其哺乳动物宿主时,细菌细胞被宿主免疫细胞吞噬。在这种情况下,φ10403S原噬菌体可以短暂切除并恢复comK基因,使细菌能够从吞噬体中逃逸。类似地,在金黄色葡萄球菌感染宿主期间,φ13的切除恢复了编码β-溶血素的基因,该基因的适时表达对发病机制至关重要。最后,在孢子形成过程中,枯草芽孢杆菌(Bacillus subtilis)母细胞中spsM基因的SPβ噬菌体切除对于将多糖添加到孢子包膜上是必要的。
总体而言,通过促进宿主的适应性和毒力,原噬菌体促进了其自身在自然界中的生存和传播。因此,原噬菌体阶段是温和噬菌体生命周期不可或缺的一部分。
重新定义"切除-复制-包装"循环
虽然处于溶原阶段的温和噬菌体可以增强宿主细胞的适应性,但裂解循环通常涉及噬菌体颗粒的产生并最终导致宿主细胞死亡。裂解循环是一个精心编排的过程,涉及在循环早期或晚期激活不同基因的表达,以有效利用细胞资源,并优化循环持续时间以产生高噬菌体滴度。
根据历史观点,原噬菌体诱导遵循一个被称为"切除-复制-包装"循环的顺序程序。在原噬菌体诱导时,方向性因子切除酶(Xis)与整合酶(Int)结合,促进原噬菌体从细菌染色体上切除。这一事件先于噬菌体裂解循环进展到后续步骤,即噬菌体DNA复制和将噬菌体DNA包装进衣壳。
这一观点基于一个前提,即噬菌体是寄生虫,它们以牺牲宿主细菌为代价来优化其繁殖。因此,早期切除随后进行噬菌体DNA复制被认为是确保最佳噬菌体传播的逻辑时间顺序。这种解释可能因许多表征原噬菌体生命周期的研究使用温度敏感突变体而得到强化。例如,沙门氏菌噬菌体P22的温度敏感突变体通常保持游离状态而非整合到染色体中,这是由于有缺陷的温度敏感阻遏蛋白的不完全抑制,允许在没有适当整合控制的情况下进行复制。
然而,"侧向转导"(lateral transduction)的发现挑战了这一观点。侧向转导是可能的,因为宿主中存在的一些原噬菌体拷贝在裂解循环后期才切除(图2)。在这种情况下,DNA复制和包装在原位进行,而原噬菌体仍整合在细菌染色体中。因此,原噬菌体DNA与下游细菌DNA一起被复制。
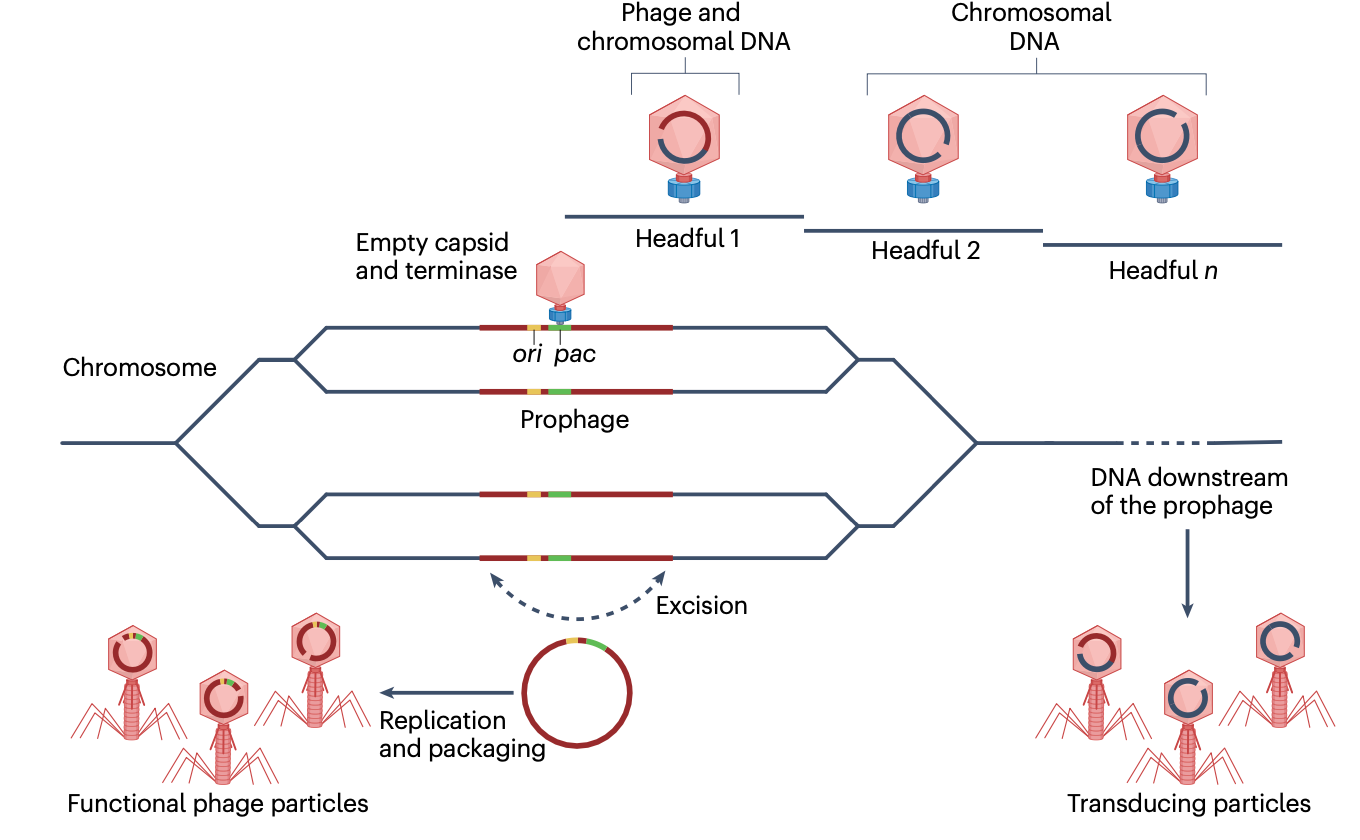
当噬菌体包装机制组装完成(发生在裂解循环后期)时,早期切除并复制的原噬菌体子集将其噬菌体基因组包装起来以产生感染性噬菌体颗粒。然而,在未切除并经历原位复制的原噬菌体子集中,包装机制识别噬菌体DNA包装信号(pac),该信号通常位于整合原噬菌体的中部。由于这些噬菌体使用"头ful包装"(headful packaging)机制来包装其DNA,在填满第一个衣壳后,包装机制将继续按照包装的方向性包装位于原噬菌体旁边的DNA。因此,该机制将继续包装原噬菌体下游的几个衣壳(头ful)的细菌DNA。结果,"切除-复制-包装"循环被重新定义,因为切除、复制和包装可以同时发生,产生带有相邻细菌染色体DNA的噬菌体颗粒,或仅带有细菌染色体DNA的颗粒,感染性噬菌体颗粒和侧向转导颗粒均在细胞裂解时释放。侧向转导的效率比普遍性转导或特异性转导高1000倍。因此,侧向转导能够以高频动员细菌染色体的大部分。
侧向转导最初在金黄色葡萄球菌噬菌体中被描述,后来也在沙门氏菌噬菌体P22和ES18、大肠杆菌噬菌体Gally、与珊瑚相关的Halomonas meridiana噬菌体Phm3以及铜绿假单胞菌噬菌体C725A中观察到。侧向转导很可能是一种普遍现象,因为原噬菌体常见于已测序的细菌基因组中,并且侧向转导仅需要延迟的原噬菌体切除和头ful包装(pac)机制。此外,在噬菌体卫星金黄色葡萄球菌致病岛(SaPIs)中也观察到了进一步的侧向转导,它们使用类似头ful包装机制的pac噬菌体。值得注意的是,cos噬菌体(如噬菌体λ)不使用头ful包装机制,也不参与侧向转导。
目前尚不清楚侧向转导如何增加噬菌体的适应性,因为它似乎减少了包装有功能性噬菌体基因组的衣壳数量。然而,侧向转导可能是一种进化适应,因为通过在诱导后更早表达切除酶可以轻易避免细菌DNA的包装。自然环境中侧向转导的证据仍然很少。尽管如此,噬菌体很可能通过侧向转导增加水平基因转移的速率来增强其潜在宿主的适应性,从而确保自身的长期生存。在金黄色葡萄球菌中,噬菌体整合位点下游的区域(可以通过侧向转导包装和转移)比上游区域更具变异性,表明进化更快。这些下游区域还包含多个致病岛和毒力基因,将侧向转导与细菌毒力联系起来。此外,观察到侧向转导可以动员金黄色葡萄球菌和大肠杆菌中染色体编码的抗噬菌体系统,这解释了染色体防御基因如何在细菌种群中轻易获得和丢失。总之,侧向转导似乎是许多温和噬菌体生命周期的一种进化而来的内在特征,它提高了水平基因转移的速率,对细菌宿主和噬菌体双方都有利。
裂解与溶原化决策的调控
当感染细菌细胞时,温和噬菌体必须要么继续进行裂解循环,要么进入溶原状态。这一决定取决于多种生理和环境因素。一旦溶原化建立,原噬菌体持续监测和整合相关输入,以决定它们是保持原噬菌体状态还是重新进入裂解循环。由于这一选择对噬菌体和细菌都至关重要,裂解与溶原化之间的决策是一个高度调控的过程。历史上,这一过程的模型是噬菌体λ的"遗传开关"。然而,以下小节描述的新发现极大地扩展了对各种噬菌体感知信号多样性的认识。这些发现强调了在决策过程中多个细菌和噬菌体之间相互作用的重要性,将生命周期调控的关注点从个体噬菌体-细菌相互作用扩展到更复杂的群落层面视角。
诱导信号的已知与未知
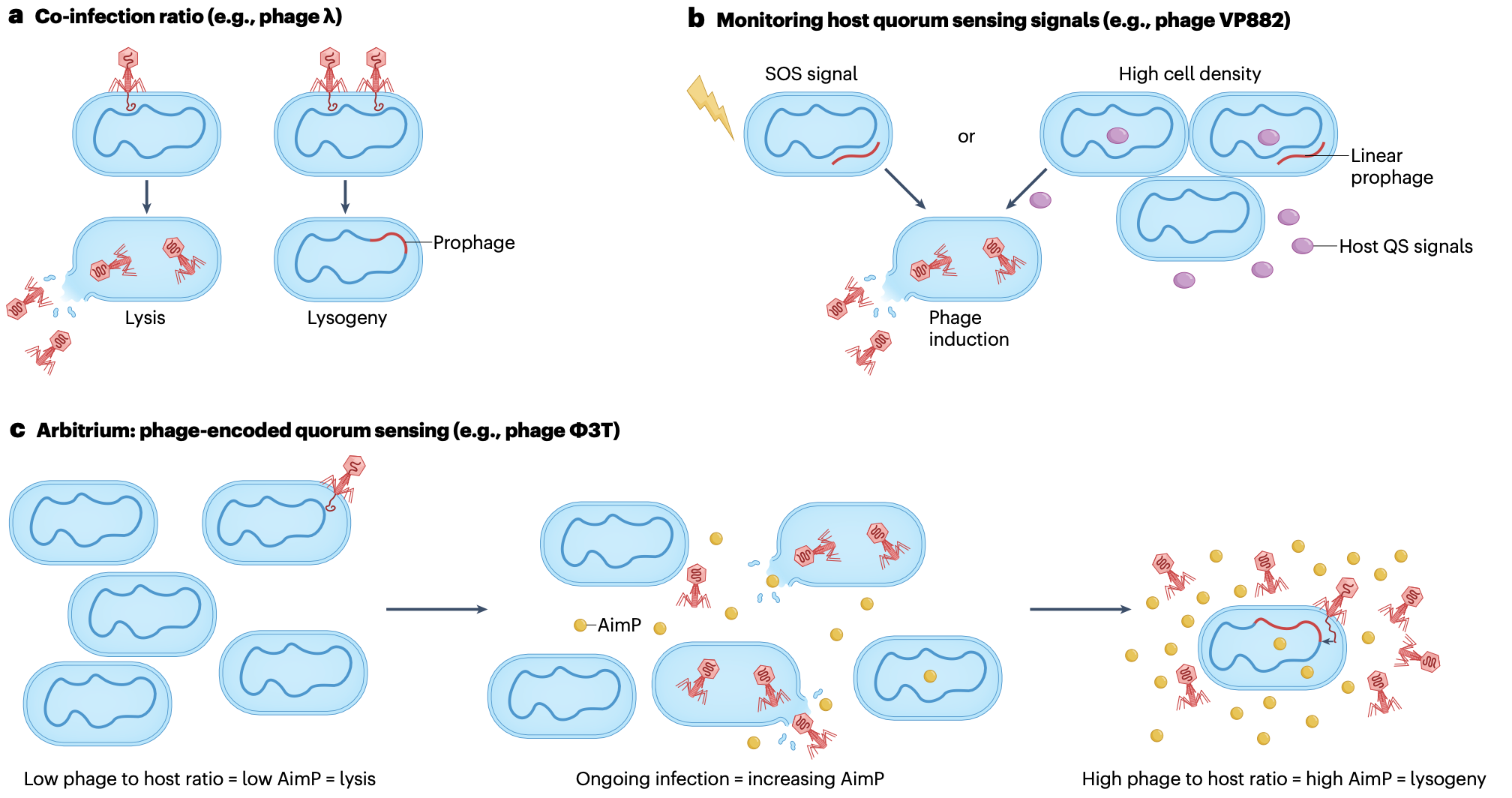
温和噬菌体从溶原化到裂解循环的转换主要在SOS反应的背景下进行研究,SOS反应是一种由DNA损伤触发的保守细菌途径。SOS反应通过刺激噬菌体主阻遏蛋白CI的自切割,导致包括λ在内的许多原噬菌体的重新激活。其他已知的需要SOS反应的原噬菌体诱导机制包括:LexA调控的小噬菌体编码抗阻遏蛋白的表达,该蛋白结合并灭活噬菌体阻遏蛋白;LexA对裂解基因的直接调控;或蛋白酶依赖性开关的激活。除了通常用于在实验室中诱导原噬菌体的DNA损伤剂(如丝裂霉素C或紫外线)外,其他条件也可以激活SOS反应,从而导致原噬菌体诱导。此类应激条件包括抗生素的存在、果糖和短链脂肪酸、氨基酸剥夺以及肠道炎症。然而,其他环境应激,如氧化应激,在某些情况下尽管激活了SOS反应,却可以抑制噬菌体产生,这说明了诱导过程如何被不同环境条件微调,并强调了理解自然环境中诱导信号的复杂性。
尽管许多原噬菌体在实验室中可以通过DNA损伤诱导,但许多噬菌体,包括那些在人类肠道中发现的噬菌体,并不编码完整的SOS诱导模块。然而,SOS反应的零星激活(例如,在复制应激期间)可能是大多数在缺乏外部诱导剂情况下在细菌种群中观察到的自发诱导事件的原因。
SOS介导的原噬菌体诱导可以在复杂微生物群落的竞争背景下被武器化。例如,colibactin(一种由几种细菌产生的代谢物,包括一些存在于肠道中的细菌)可以诱导邻近细菌中的原噬菌体,导致竞争者死亡。产生colibactin的细菌通过表达colibactin抗性蛋白来避免同样的命运,这些蛋白阻止其自身原噬菌体的诱导。类似地,肺炎链球菌(Streptococcus pneumoniae)通过产生H2O2来根除竞争细菌,H2O2会致命地激活邻近溶原性金黄色葡萄球菌细胞中的原噬菌体诱导,但不会激活肺炎链球菌菌株。
一些原噬菌体的诱导是由不激活SOS反应的信号触发的。例如,在大肠杆菌中,盐胁迫诱导噬菌体434,阳离子螯合诱导噬菌体933W,特定宿主基因的过表达可以诱导噬菌体21、Φ80和434。此外,一些原噬菌体编码由细菌群体感应信号激活的诱导模块,表明噬菌体诱导可以受群落相互作用的调控。
尽管进行了广泛的研究,但对温和噬菌体中诱导信号的多样性仍知之甚少。例如,感染人类肠道共生菌Bacteroides thetaiotaomicron的Mushu噬菌体和大肠杆菌噬菌体P2即使在某些条件下产生功能性病毒粒子,也不响应经典的诱导信号。此外,对肠道细菌、清酒乳杆菌(Lactilactobacillus sakei)和环境枯草芽孢杆菌溶原菌的筛选显示,尽管这些细菌基因组中存在许多预测的原噬菌体,但只有一部分测试菌株在暴露于丝裂霉素C或紫外线时发生裂解。重要的是,对于一些对丝裂霉素C无诱导反应的肠道细菌原噬菌体,诱导可以通过其他条件触发,包括碳耗竭、甜菊糖的存在或通过未知机制的人类宿主相关细胞产物的存在。
一些温和噬菌体对多种不同诱导信号作出反应的能力可能有助于在特定环境和生态位中调节裂解与溶原化之间的决策。例如,在多重溶原菌(polylysogens)中,多种诱导途径可能是有利的。作为对共同触发因素(如SOS反应)的响应,多个共存的噬菌体可以被诱导并竞争资源。相比之下,如果诱导信号是噬菌体特异性的,则主要产生一种特定的噬菌体。例如,在金黄色葡萄球菌菌株中,只有噬菌体MBL3对铜绿假单胞菌产生的代谢物绿脓菌素(phenazine pyocyanin)敏感。噬菌体特异性诱导信号可以防止多重溶原菌中所有原噬菌体的同时诱导,从而避免资源竞争。
监测和利用宿主群体感应
除了响应环境应激源外,一些温和噬菌体还监测细菌群体感应信号,以评估宿主细胞密度并相应地调整其生命周期决策。例如,铜绿假单胞菌噬菌体D3112和JBD30的裂解-溶原化决策受细菌群体感应分子N-酰基高丝氨酸内酯(AHLs)的调控。在高AHL浓度(指示高细胞密度)下,这些噬菌体倾向于溶原化,而在低AHL浓度(指示低细胞密度)下,它们倾向于裂解循环。这种调节被认为可以优化噬菌体传播,因为在低细胞密度下,裂解循环可能更有利,因为新宿主稀缺,而溶原化可以确保在宿主种群中持续存在。
此外,一些枯草芽孢杆菌温和噬菌体,如Φ3T,编码自己的群体感应系统,称为"仲裁系统"(arbitrium),用于估计噬菌体与宿主的比例。在感染期间,噬菌体产生通信肽AimP,该肽被释放到环境中并被邻近细胞导入。在感染早期,AimP浓度低,感染噬菌体倾向于裂解循环。然而,高AimP水平表明噬菌体与宿主比例高,则倾向于溶原化,因为找到未来宿主感染的概率较低。仲裁系统使噬菌体能够根据局部噬菌体密度做出集体决策,防止过度捕食和宿主种群的崩溃。
这些例子表明,温和噬菌体不仅响应宿主的生理状态,还积极监测和利用细菌的通信系统,甚至发展出自己的通信机制,以优化其生命周期策略。这种复杂的调控网络强调了噬菌体-宿主相互作用的动态和群落层面性质。
规避细胞防御
细菌已经进化出多种防御系统来对抗噬菌体感染,包括限制修饰系统、CRISPR-Cas系统和多种流产感染系统。为了成功繁殖,噬菌体必须规避或克服这些防御机制。温和噬菌体,特别是原噬菌体,在帮助宿主抵抗其他噬菌体感染方面发挥着独特作用,但它们自身在感染新宿主时也必须应对外部防御。
原噬菌体编码的抗噬菌体防御系统可以保护溶原菌免受超感染(superinfection)。例如,许多原噬菌体携带编码毒素-抗毒素系统、限制修饰系统或CRISPR-Cas变体的基因,这些系统可以靶向并消除入侵的竞争噬菌体。这种"以毒攻毒"的策略确保了原噬菌体在宿主内的独占性,并减少了资源竞争。
然而,当温和噬菌体从溶原菌中诱导并试图感染新宿主时,它可能遇到该宿主编码的防御系统。为了应对这一挑战,一些温和噬菌体进化出了对抗特定防御系统的机制。例如,T4噬菌体编码蛋白质可以抑制大肠杆菌的限制修饰系统。类似地,一些噬菌体具有抗CRISPR(Acr)蛋白,可以阻断CRISPR-Cas系统的活性。
此外,温和噬菌体可能通过改变其基因组序列或包装机制来逃避宿主防御。例如,通过突变其基因组中限制性酶切位点或使用化学修饰(如羟甲基胞嘧啶)来保护其DNA免受限制性内切酶切割。一些噬菌体还使用蛋白质"盔甲"来保护其DNA在注入过程中不被宿主核酸酶降解。
规避细胞防御的能力是温和噬菌体成功感染和传播的关键。这种能力不仅影响个体感染事件的结果,还在塑造微生物群落的组成和进化动态中发挥重要作用。
防止噬菌体携带防御系统的自身免疫
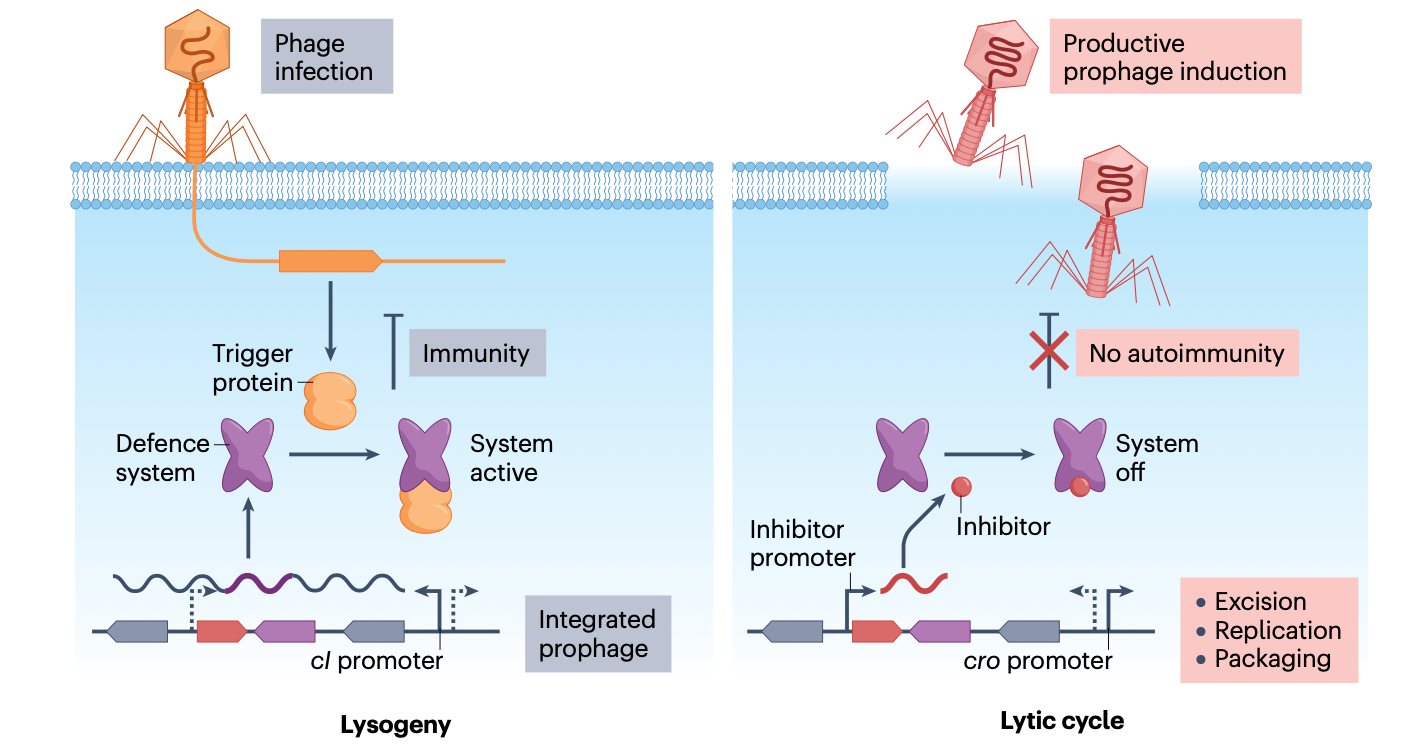
当原噬菌体编码抗噬菌体防御系统时,它们面临着一个独特的挑战:如何防止这些系统攻击原噬菌体自身或同一细胞内的其他原噬菌体?这种自身免疫(autoimmunity)风险需要精细的调控机制来确保防御系统仅在适当的时候被激活。
一种常见的策略是时空隔离。例如,原噬菌体编码的防御系统可能仅在裂解循环期间表达,而在溶原状态下保持沉默。这样,当原噬菌体处于整合状态时,防御系统不会攻击宿主染色体或原噬菌体自身。另一种策略是使用特异性调节蛋白,确保防御系统仅靶向外来噬菌体,而不识别原噬菌体自身的DNA。
例如,一些原噬菌体编码的毒素-抗毒素系统在溶原状态下被紧密调控,只有在特定诱导信号(如DNA损伤)下才会被激活。类似地,原噬菌体携带的CRISPR-Cas系统可能被编程为不靶向原噬菌体自身的序列,或者通过表达抗CRISPR蛋白在溶原状态下自我抑制。
防止自身免疫对于维持原噬菌体在宿主内的稳定共存至关重要。失败可能导致原噬菌体自身被消除或宿主细胞死亡,这对噬菌体和宿主都不利。因此,温和噬菌体进化出了复杂的调控网络来平衡防御效益和自身免疫风险。
温和噬菌体与其他可移动遗传元件的相互作用
温和噬菌体并非孤立存在;它们与细菌基因组中的其他可移动遗传元件(MGEs)相互作用,包括质粒、转座子、整合性接合元件和噬菌体卫星(如致病岛)。这些相互作用可以是竞争性的,也可以是合作性的,共同塑造了细菌的基因组可塑性和进化。
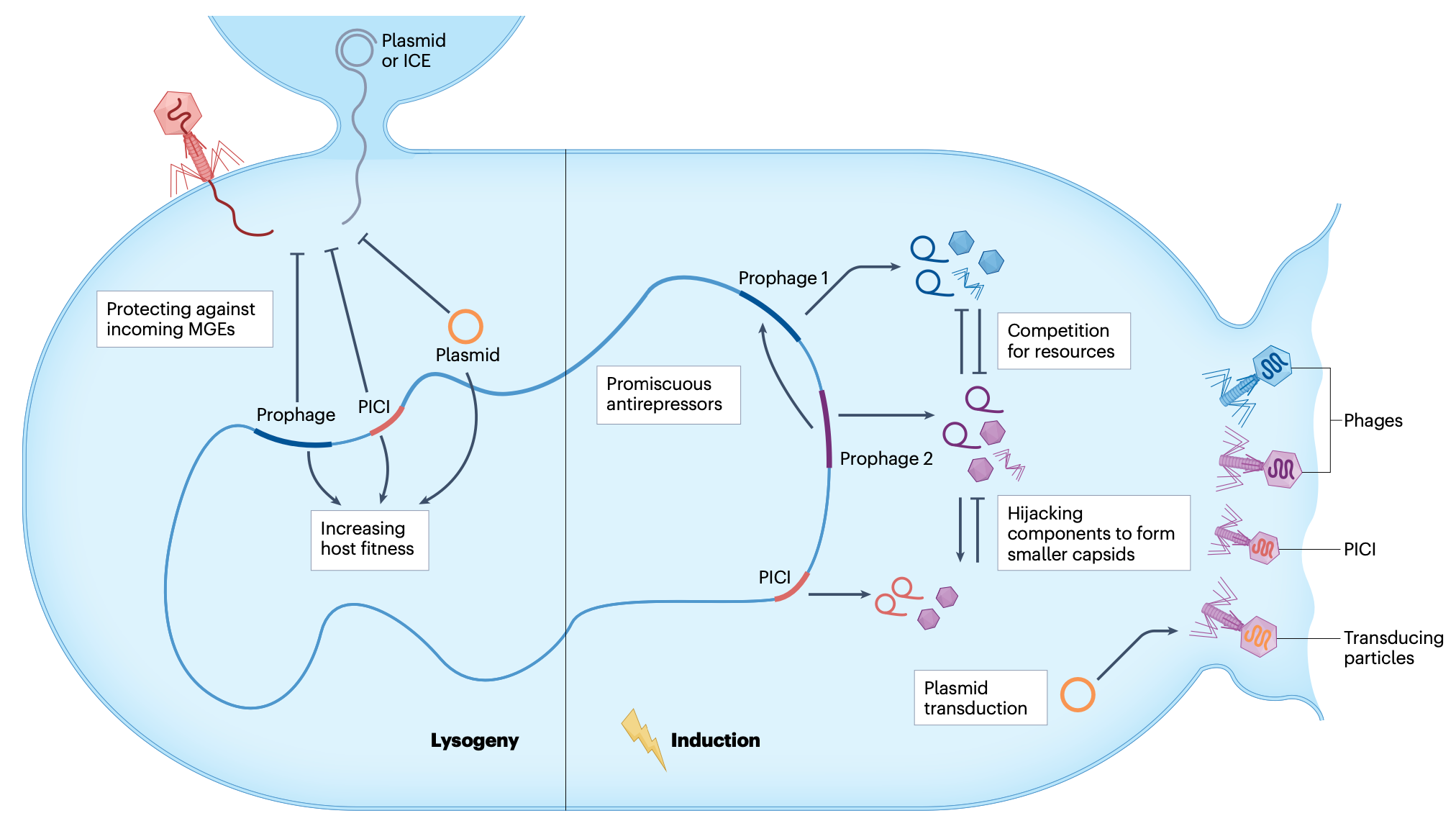
与质粒的相互作用
质粒和温和噬菌体经常在细菌细胞内共存,它们可能竞争复制资源或协同工作以促进基因转移。例如,一些质粒编码的因子可以抑制原噬菌体的诱导,从而维持溶原状态。相反,原噬菌体诱导有时可以动员质粒的接合转移,通过细胞裂解促进质粒传播到邻近细胞。
此外,质粒和原噬菌体可以交换遗传模块。质粒可能获得原噬菌体的整合酶或切除酶基因,从而获得整合能力。同样,原噬菌体可能从质粒获得抗生素抗性基因或毒力因子,增强其宿主的适应性。
与噬菌体卫星的相互作用
噬菌体卫星,如金黄色葡萄球菌致病岛(SaPIs),是依赖辅助噬菌体进行包装和传播的可移动遗传元件。SaPIs通常整合在宿主染色体中,当辅助噬菌体诱导时,它们可以被动员并包装进噬菌体样颗粒中。有趣的是,SaPIs可以干扰辅助噬菌体的生命周期,例如通过劫持其包装机制或抑制其复制,从而确保自身的优先传播。
温和噬菌体与SaPIs之间的这种相互作用是一种"寄生虫中的寄生虫"关系,其中SaPIs利用噬菌体的机制进行传播,同时调节噬菌体的活性以最大化自身利益。这种动态关系可以影响毒力基因的传播和细菌群落的进化。
原噬菌体之间的相互作用
在多重溶原菌中,多个原噬菌体可能共存于同一细菌基因组中。这些原噬菌体可以相互竞争或合作。例如,一个原噬菌体的诱导可能触发其他原噬菌体的协同诱导,导致"裂解爆发"(lysis burst)。相反,一些原噬菌体编码的阻遏蛋白可以交叉抑制其他原噬菌体的诱导,维持溶原状态的稳定。
原噬菌体之间的遗传交换也很常见,通过同源重组或转座事件产生新的噬菌体变体。这种遗传混合可以产生具有新宿主范围或毒力特性的噬菌体,促进细菌种群的快速进化。
总之,温和噬菌体与其他可移动遗传元件的相互作用增加了细菌基因组可塑性的另一层复杂性。这些相互作用不仅影响个体噬菌体和元件的命运,还在水平基因转移、抗生素抗性传播和毒力进化中发挥关键作用。
结论
温和噬菌体的生命周期远不止是裂解和溶原化之间的简单切换。本综述整合了最新研究进展,提出了一个更为全面和动态的生命周期框架,其中原噬菌体阶段、与宿主防御系统的相互作用、与其他可移动遗传元件的竞争与合作,以及群落层面的信号整合都是不可或缺的组成部分。
原噬菌体并非被动乘客,而是通过表达附属基因主动塑造宿主生理和适应性,在溶原期间提供抗噬菌体防御,并通过"活性溶原"机制充当调控开关。裂解循环的经典"切除-复制-包装"范式已被"侧向转导"等发现所扩展,揭示了噬菌体如何以高频动员细菌染色体片段,从而成为水平基因转移的强大引擎。
裂解与溶原化的决策是一个高度复杂的过程,整合了来自宿主生理、环境压力、群体感应甚至噬菌体自身通信系统的多种信号。这种多层次的调控使噬菌体能够优化其传播策略,响应不断变化的环境条件。
此外,温和噬菌体与细菌防御系统以及其他可移动遗传元件的相互作用凸显了微生物世界中相互关联的动态网络。噬菌体不仅是细菌的寄生虫,也是细菌进化、适应性和群落生态的关键驱动因素。
未来研究需要进一步探索温和噬菌体在自然环境和宿主相关微生物组中的多样性和功能。了解这些复杂相互作用的分子机制和生态后果,将有助于开发新的噬菌体疗法、操纵微生物群落,并应对抗生素耐药性等全球健康挑战。最终,将温和噬菌体视为具有复杂生命周期和广泛生态影响的动态实体,将丰富我们对微生物世界及其与人类健康和疾病关系的理解。