Single-cell analyses identify the ginseng embryonic protoderm as a native compartment for high-efficiency ginsenoside production
单细胞测序分析证实:人参胚胎原表皮是高效合成人参皂苷的天然功能分区

摘要
人参的药用价值主要来源于其次级代谢产物人参皂苷 。但人参栽培周期漫长、易受病害侵染,严重制约了人参产业的发展。本研究证实,人参胚胎原表皮具备高效合成人参皂苷的能力。结合单细胞转录组与质谱成像分析发现,人参胚胎愈伤组织中原表皮的人参皂苷积累量可与林下参相媲美。表观遗传研究表明,胚胎愈伤组织中再生通路及人参皂苷代谢通路相关基因位点的组蛋白乙酰化水平升高、染色质开放程度增强 ,与其皂苷合成能力密切相关。提升组蛋白乙酰化水平,或过表达植株再生关键基因WOX11 ,均可进一步显著促进胚胎愈伤组织合成人参皂苷。综上,胚胎愈伤组织原表皮可作为高效合成人参皂苷的天然生物功能区域,为人参皂苷量产提供了有别于传统田间栽培的新型技术途径。
引言
植物天然产物是医药原料、食品添加剂与新型生物燃料的重要来源,已然成为细胞工厂领域的研究热点。优化植物天然产物的生物合成制备技术,不仅能够保护珍稀药用植物资源、降低生产成本,更是推动产业绿色可持续发展的关键举措。
人参皂苷主要来源于人参,依据化学结构可分为三大类,即原人参二醇型、原人参三醇型与齐墩果烷型,各类皂苷均具备多样的药理活性。其中,人参皂苷 Ro 具有抗炎、减脂功效;竹节参皂苷 IVa 能够保护骨骼、延缓衰老;人参皂苷 Re 可发挥降糖、神经调控与抗肿瘤作用;人参皂苷 Rh1 则兼具免疫调节、抗氧化及心血管保护等活性。凭借丰富的药用价值,人参成为高价值经济作物,全球市场需求量逐年攀升。
然而人参产业发展面临诸多现实难题:其种植采收周期长达 4 至 25 年,培育条件严苛,还存在连作障碍、易受病原菌侵染及逆境胁迫等问题,极大限制了产业规模化发展。为此,兼具环保性与经济性的人参细胞培养技术已逐步应用于食品、美妆、医药等多个领域。但该技术仍存在明显短板,主要表现为细胞再生能力弱、人参皂苷等次生代谢产物合成量偏低,难以实现工业化量产。
植物体内多数次生代谢产物存在细胞特异性分区积累特征,这也是植物抵御内源代谢产物毒性的重要自我保护机制。明确次生代谢产物富集的特异细胞类型,有助于解析其合成调控通路,进而提升人参悬浮培养体系中皂苷的合成效率。已有研究证实,人参皂苷的生物合成存在多细胞分区合成特点,但参与该过程的原始功能细胞类型至今尚不明确。
体细胞胚胎发生是植物体细胞分化形成类胚胎结构的无性发育途径,依托该体系结合基因编辑技术,可高效提升植物天然产物合成积累量。人参胚性愈伤组织自 1980 年成功建立以来,被证实兼具优异的植株再生能力与人参皂苷合成能力,不过目前关于人参胚性愈伤组织的再生调控机制,及其药用活性代谢物合成的分子机理仍缺乏系统性解析。
模式植物拟南芥相关研究表明,生长素可通过重塑染色质开放状态快速重编程细胞全能性通路,进而启动体细胞胚胎发生进程;在此过程中,多条生长素通路核心转录因子协同调控植株体细胞胚胎发育,主要包括AGL15 、FUS3 、LEC1/2 、ABI3 、BBM 、PLT 家族、WUS 、WOX 家族、LBD40 、IAA30 与YUCCA 等基因。此外,以组蛋白乙酰化为代表的表观遗传修饰,可通过调控关键功能基因的转录表达,深度参与体细胞胚胎发生进程。
本研究以人参叶片为材料,分别诱导获得胚性愈伤组织与非胚性愈伤组织,二者在再生能力与人参皂苷积累模式上存在显著差异。借助单细胞测序与多组学联合分析,我们探明了细胞重编程过程中人参皂苷生物合成的表观遗传调控机制,为人参新型组培体系构建及皂苷高效合成优化奠定了重要分子理论基础。
结果
人参胚性愈伤组织可高效合成人参皂苷
为验证愈伤组织作为体外合成人参皂苷培养材料的可行性,本研究将人参叶片接种至添加 2 mg/L 萘乙酸与 0.5 mg/L 6 - 苄氨基嘌呤的 MS 培养基中,成功诱导获得叶片来源愈伤组织。后续将该愈伤组织置于两种不同培养体系中继代培养: 其一,在添加 2.0 mg/L 2,4 - 二氯苯氧乙酸与 0.5 mg/L 激动素的 MS 培养基中培养,获得质地疏松、呈半透明白色、无芽分化再生能力的非胚性愈伤组织; 其二,将愈伤组织转接至无外源激素的 MS 培养基中连续培养 5 个月,诱导得到质地紧实、色泽偏黄且可持续分化形成体细胞胚的胚性愈伤组织。将胚性愈伤组织诱导形成的体细胞胚置于含 0.1 mg/L 萘乙酸与 0.5 mg/L 激动素的培养基中培养,可进一步分化形成不定芽。
采用特异性染色试剂苏丹红 7B 对两类愈伤组织进行染色鉴定,该试剂可特异性标记胚胎组织中富集的甘油三酯。染色结果显示,胚性愈伤组织可观察到明显呈红色的类胚胎结构,而非胚性愈伤组织无此类结构,染色后整体呈白色。
本研究以林下参和园栽人参作为对照,系统测定两类愈伤组织的人参皂苷合成能力。超高效液相色谱 - 串联质谱检测结果表明,胚性愈伤组织的人参皂苷总含量与林下参相近,显著高于园栽人参与非胚性愈伤组织。 在单体皂苷组分上,胚性愈伤组织中人参皂苷 Re、齐墩果烷型皂苷(Ro、竹节参皂苷 IVa)及稀有皂苷(Rg2、Rh1、F2、F1)的积累量不仅远超园栽人参与非胚性愈伤组织,部分组分含量甚至优于林下参。
多糖含量测定结果显示,胚性愈伤组织多糖含量显著高于非胚性愈伤组织,但低于林下参和园栽人参;而其总黄酮含量则显著高于其余所有试验材料。 上述试验结果证实,人参胚性愈伤组织可作为优质体外培养材料,能够高效合成与人参植株同源的药用活性皂苷,具备极高的体外量产应用潜力。
人参皂苷在胚胎原表皮中合成
为阐明胚性愈伤组织可高效合成人参皂苷、而非胚性愈伤组织不具备该能力的分子与细胞机制,本研究对两类愈伤组织开展了转录组测序与单细胞转录组测序分析。
转录组结果显示,大量植株再生与人参皂苷代谢通路相关基因在胚性愈伤组织中显著富集,在非胚性愈伤组织中则表达水平极低。 其中,胚性愈伤组织中富集的再生相关基因主要包括:AP2 类转录因子家族中与分生组织发育相关的 euANT 分支基因(如PLT 、ANT 、BBM )、干细胞调控相关WOX 家族基因(如WOX11A 、WOX11B )、胚胎发育关键调控基因(如ABI3B 、LEC1A/B 、AGL15A )、LBD 家族基因(如LBD16A 、LBD16B )以及生长素合成通路基因(如YUCCA 、TAA1 )。 富集的皂苷代谢相关基因则涵盖齐墩果烷型、原人参二醇型、原人参三醇型皂苷合成通路关键基因,包括鲨烯环氧酶、β- 香树素合成酶、达玛烯二醇 - II 合成酶及多种CYP450 家族功能基因。
单细胞转录组测序将两类愈伤组织细胞划分为 0~9 共 9 个细胞类群。 利用标志基因完成细胞类型注释:0 类群表达基因数量少且无特异标志基因为未知细胞;1 类群表达FAD2 ,判定为基本组织细胞;2 类群特异性表达ATML1 与PDF2 ,为原表皮细胞 ;3~5 类群表达PLT3B 、WOX11 、ABI3B 等基因,属于胚胎分生组织细胞;6 类群高表达TUA2 、CKS2 ,为 G2/M 期分裂细胞;7 类群富集组蛋白基因,代表 S 期增殖细胞;8、9 类群细胞类型暂未明确。
原位杂交试验进一步验证,PLT3B 、AIL1 等再生核心基因在胚胎维管前体细胞群与子叶胚胎细胞群中存在特异表达。 细胞丰度差异分析结果表明,非胚性愈伤组织中缺失 2~5 号细胞类群,说明其几乎不含原表皮细胞与胚胎分生组织细胞。 与此同时,人参皂苷合成通路关键基因均在胚性愈伤组织的 2 号原表皮细胞类群中高度特异表达。
对β-AS1 基因开展原位杂交定位发现,该基因在胚性愈伤组织及体细胞胚的原表皮中大量表达。常压微探针基质辅助激光解吸电离质谱成像结果证实,人参皂苷 Ro、Rd 主要富集于胚性愈伤组织与体细胞胚的原表皮部位。
上述结果证实,胚性愈伤组织的原表皮是合成人参皂苷的天然特异功能区域 。
染色质开放程度提升保障人参皂苷合成
为揭示胚性愈伤组织合成人参皂苷的内在分子机制,本研究对比分析了胚性与非胚性愈伤组织的染色质状态。 蛋白免疫印迹结果表明,作为基因激活表达标志的组蛋白 H3K9 乙酰化修饰在胚性愈伤组织中水平较高,在非胚性愈伤组织中几乎检测不到;而同样参与基因表达调控的 H3K4me3、H3K4me2 修饰水平在两类材料中无明显差异。
染色质免疫共沉淀测序结果显示,胚性愈伤组织中全基因组范围的 H3K9 乙酰化修饰大量富集于基因转录起始位点附近,非胚性愈伤组织无此特征;且皂苷合成关键基因与植株再生相关基因位点均存在高水平 H3K9 乙酰化修饰。
组蛋白乙酰化修饰能够改变染色质紧致程度,因此本研究借助转座酶可及染色质测序技术检测两类愈伤组织的染色质开放水平。结果显示,胚性愈伤组织整体染色质开放度显著高于非胚性愈伤组织,且绝大多数人参皂苷代谢基因、体细胞胚胎再生相关基因均呈现更高的染色质可及性。
综上可知,相较于非胚性愈伤组织,胚性愈伤组织中皂苷合成与植株再生相关基因位点具备更高的组蛋白乙酰化水平与染色质开放程度,这是其维持胚胎细胞特性、顺利启动人参皂苷代谢合成的重要染色质基础。
胚胎原表皮可作为人参皂苷原位生物合成功能区
基于染色质修饰与再生调控基因的相关研究结论,本研究通过人工调控手段,靶向改造胚性愈伤组织胚胎原表皮的生理状态,以期进一步提升人参皂苷合成效率。
首先分别采用组蛋白乙酰转移酶抑制剂姜黄素、组蛋白去乙酰化酶抑制剂曲古抑菌素 A 处理两类愈伤组织。处理十天后发现,曲古抑菌素 A 可显著上调皂苷合成通路基因表达,促进人参皂苷积累;而姜黄素则会抑制相关基因表达、降低皂苷合成量,直接证实组蛋白乙酰化水平是调控人参皂苷合成的核心关键因素。该调控规律在三七中同样成立,说明该机制在人参属植物中具有保守性。
其次,本研究在胚性愈伤组织中过表达WOX11B 基因以强化其再生能力。结果显示,过表达WOX11B 能够显著提升胚性愈伤组织再生效率与遗传转化效率,有效诱导发根发生;同时可显著上调皂苷代谢通路基因表达,大幅提升皂苷合成积累能力。
以上试验表明,施用曲古抑菌素 A 或过表达WOX11B 基因,均可进一步强化胚胎原表皮的皂苷合成能力,是提升体外体系人参皂苷产量的有效手段。
人参皂苷在胚胎原表皮中合成
为阐明胚性愈伤组织可高效合成人参皂苷、而非胚性愈伤组织不具备该能力的分子与细胞机制,本研究对两类愈伤组织开展了转录组测序与单细胞转录组测序分析。
转录组结果显示,大量植株再生与人参皂苷代谢通路相关基因在胚性愈伤组织中显著富集,在非胚性愈伤组织中则表达水平极低。 其中,胚性愈伤组织中富集的再生相关基因主要包括:AP2 类转录因子家族中与分生组织发育相关的 euANT 分支基因(如PLT 、ANT 、BBM )、干细胞调控相关WOX 家族基因(如WOX11A 、WOX11B )、胚胎发育关键调控基因(如ABI3B 、LEC1A/B 、AGL15A )、LBD 家族基因(如LBD16A 、LBD16B )以及生长素合成通路基因(如YUCCA 、TAA1 )。 富集的皂苷代谢相关基因则涵盖齐墩果烷型、原人参二醇型、原人参三醇型皂苷合成通路关键基因,包括鲨烯环氧酶、β- 香树素合成酶、达玛烯二醇 - II 合成酶及多种CYP450 家族功能基因。
单细胞转录组测序将两类愈伤组织细胞划分为 0~9 共 9 个细胞类群。 利用标志基因完成细胞类型注释:0 类群表达基因数量少且无特异标志基因为未知细胞;1 类群表达FAD2 ,判定为基本组织细胞;2 类群特异性表达ATML1 与PDF2 ,为原表皮细胞 ;3~5 类群表达PLT3B 、WOX11 、ABI3B 等基因,属于胚胎分生组织细胞;6 类群高表达TUA2 、CKS2 ,为 G2/M 期分裂细胞;7 类群富集组蛋白基因,代表 S 期增殖细胞;8、9 类群细胞类型暂未明确。
原位杂交试验进一步验证,PLT3B 、AIL1 等再生核心基因在胚胎维管前体细胞群与子叶胚胎细胞群中存在特异表达。 细胞丰度差异分析结果表明,非胚性愈伤组织中缺失 2~5 号细胞类群,说明其几乎不含原表皮细胞与胚胎分生组织细胞。 与此同时,人参皂苷合成通路关键基因均在胚性愈伤组织的 2 号原表皮细胞类群中高度特异表达。
对β-AS1 基因开展原位杂交定位发现,该基因在胚性愈伤组织及体细胞胚的原表皮中大量表达。常压微探针基质辅助激光解吸电离质谱成像结果证实,人参皂苷 Ro、Rd 主要富集于胚性愈伤组织与体细胞胚的原表皮部位。
上述结果证实,胚性愈伤组织的原表皮是合成人参皂苷的天然特异功能区域 。
染色质开放程度提升保障人参皂苷合成
为揭示胚性愈伤组织合成人参皂苷的内在分子机制,本研究对比分析了胚性与非胚性愈伤组织的染色质状态。 蛋白免疫印迹结果表明,作为基因激活表达标志的组蛋白 H3K9 乙酰化修饰在胚性愈伤组织中水平较高,在非胚性愈伤组织中几乎检测不到;而同样参与基因表达调控的 H3K4me3、H3K4me2 修饰水平在两类材料中无明显差异。
染色质免疫共沉淀测序结果显示,胚性愈伤组织中全基因组范围的 H3K9 乙酰化修饰大量富集于基因转录起始位点附近,非胚性愈伤组织无此特征;且皂苷合成关键基因与植株再生相关基因位点均存在高水平 H3K9 乙酰化修饰。
组蛋白乙酰化修饰能够改变染色质紧致程度,因此本研究借助转座酶可及染色质测序技术检测两类愈伤组织的染色质开放水平。结果显示,胚性愈伤组织整体染色质开放度显著高于非胚性愈伤组织,且绝大多数人参皂苷代谢基因、体细胞胚胎再生相关基因均呈现更高的染色质可及性。
综上可知,相较于非胚性愈伤组织,胚性愈伤组织中皂苷合成与植株再生相关基因位点具备更高的组蛋白乙酰化水平与染色质开放程度,这是其维持胚胎细胞特性、顺利启动人参皂苷代谢合成的重要染色质基础。
胚胎原表皮可作为人参皂苷原位生物合成功能区
基于染色质修饰与再生调控基因的相关研究结论,本研究通过人工调控手段,靶向改造胚性愈伤组织胚胎原表皮的生理状态,以期进一步提升人参皂苷合成效率。
首先分别采用组蛋白乙酰转移酶抑制剂姜黄素、组蛋白去乙酰化酶抑制剂曲古抑菌素 A 处理两类愈伤组织。处理十天后发现,曲古抑菌素 A 可显著上调皂苷合成通路基因表达,促进人参皂苷积累;而姜黄素则会抑制相关基因表达、降低皂苷合成量,直接证实组蛋白乙酰化水平是调控人参皂苷合成的核心关键因素。该调控规律在三七中同样成立,说明该机制在人参属植物中具有保守性。
其次,本研究在胚性愈伤组织中过表达WOX11B 基因以强化其再生能力。结果显示,过表达WOX11B 能够显著提升胚性愈伤组织再生效率与遗传转化效率,有效诱导发根发生;同时可显著上调皂苷代谢通路基因表达,大幅提升皂苷合成积累能力。
以上试验表明,施用曲古抑菌素 A 或过表达WOX11B 基因,均可进一步强化胚胎原表皮的皂苷合成能力,是提升体外体系人参皂苷产量的有效手段。
讨论
目前大量药物制剂、膳食保健品、食品及化妆品中均添加植物源天然活性成分。植物细胞工厂凭借经济高效、绿色环保与可持续等优势,已成为规模化制备各类高价值天然产物的重要技术路径。相较于传统田间种植模式,植物细胞工厂前期投入成本偏高,但依托生物反应器可实现生物量高效积累,且生产过程不受季节气候限制,因此在次生代谢产物规模化生产领域具备广阔商业化应用前景。而传统田间栽培模式易受气候波动、病原菌侵染等外界不确定因素制约,生产稳定性较差。
本研究借助单细胞测序技术证实,人参胚胎原表皮可作为合成人参皂苷的天然特异功能细胞区域。胚胎原表皮细胞富含脂质及其衍生物,十分适宜合成人参皂苷 Ro、竹节参皂苷 IVa 等脂溶性植物次生代谢产物。据此推测,胚胎原表皮不仅能在人参中高效合成人参皂苷,在其他植物中也有望作为合成各类脂溶性次生代谢物的理想天然合成位点。已有研究证实人参植株体内人参皂苷存在多细胞分区合成特征,本研究结果也进一步佐证了该类活性物质具有特定细胞类型定向合成的特性。
染色质状态重塑是体细胞重编程并进入胚胎发育状态的核心关键环节。本研究证实,人参胚性愈伤组织的形成与全基因组组蛋白乙酰化修饰水平上调密切相关。组蛋白乙酰化水平升高能够显著提升染色质开放程度,进而解除胚胎发育与人参皂苷代谢通路相关基因的转录抑制,启动相关基因正常表达。因此,利用曲古抑菌素 A 等组蛋白去乙酰化酶抑制剂提升组蛋白乙酰化水平,是强化胚胎原表皮皂苷合成功能简便且有效的调控手段。
调控植株再生相关基因同样能够优化胚胎原表皮的皂苷合成效能。本研究证实,过表达WOX11 基因既可提升人参胚性愈伤组织再生能力,还能显著促进人参皂苷代谢合成通路运转。除WOX11 外,PLT 、LEC 等诸多再生调控基因也具备作为分子工具、优化原表皮皂苷合成效率的潜力。后续可进一步探究多基因协同调控组合,并在不同植物物种中验证其调控效果。
综上,联合运用染色质表观调控、再生核心基因过表达,结合脂类代谢通路基因的遗传转化与基因编辑技术,有望在不同植物物种中改造优化胚胎原表皮功能,搭建可高效合成各类植物脂溶性活性代谢物的新型合成体系(图 6)。
材料与方法
植物材料
以三年生吉林人参叶片诱导获得初代愈伤组织,接种至含 2 mg/L 萘乙酸、0.5 mg/L 6 - 苄氨基嘌呤、3% 蔗糖的 MS 培养基中,置于 25±1℃黑暗条件下培养。将初代愈伤组织转接至添加 1.0 mg/L 2,4 - 二氯苯氧乙酸与 0.5 mg/L 激动素的培养基中,相同条件下继代培养 10 代以上(每 5 周继代一次),获得非胚性愈伤组织;叶片愈伤组织在无外源激素 MS 培养基中连续培养 5 个月,诱导得到胚性愈伤组织。所有试验材料取样后立即液氮速冻,置于 - 80℃冰箱保存。代谢组与转录组试验设置 3 组生物学重复,单细胞转录组、ATAC-seq 及 ChIP-seq 试验均设置 2 组生物学重复。
芽再生能力测定
将胚性愈伤组织置于不含激素的固体 MS 培养基(含维生素、8 g/L 琼脂,pH 5.8)黑暗培养;非胚性愈伤组织先接种于添加 1 mg/L 2,4-D 与 0.5 mg/L 激动素的 MS 培养基中黑暗预培养一周,随后将两类愈伤组织一同转接至芽分化诱导培养基(含 0.1 mg/L 萘乙酸、0.5 mg/L 激动素、20 g/L 蔗糖、8 g/L 琼脂,pH 5.8)培养 1 个月。统计外植体总数与再生芽数量,计算芽再生率(再生芽数 / 总外植体数),试验共计设置 26 组生物学重复。
人参皂苷代谢组检测
参照已建立的检测方法开展人参皂苷代谢组定量分析。采用 Waters 超高效液相色谱仪搭配 HSS T3 色谱柱(2.1×100 mm,1.8 μm),柱温 40℃,流速 0.4 mL/min;流动相 A 为 0.1% 甲酸 - 乙腈溶液,流动相 B 为 0.1% 甲酸 - 水溶液,设置梯度洗脱程序:0~2.5 min,A 相占比 28%~31%;2.5~3 min,31%~35%;3~4.5 min,35%~36%;4.5~6 min,36%~37%;6~7 min,37%~50%;7~9 min,50%~98%;9~10 min,维持 98%;10~11 min,回落至 28%;11~13 min,28% 平衡色谱柱,进样体积 1 μL。采用负离子扫描模式,优化设置一级质谱、二级质谱、去簇电压、碰撞能、碰撞室出口电压等参数。
取 20~30 mg 冻干样品,加入 1.5 mL 75% 甲醇溶液浸泡过夜,500 W、40 kHz 条件下超声提取 30 min;13000 rpm、20℃离心 10 min,上清液经 0.22 μm 滤膜过滤后 4℃保存,用于超高效液相色谱 - 串联质谱联用检测。配制 24 种人参皂苷标准品母液,稀释配制浓度为 20 μg/mL 的混合工作液,4℃冷藏备用。
采用 Waters MassLynx 4.1 软件采集质谱数据,AB SCIEX MultiQuant 软件完成峰识别与定量分析,参数设置:保留时间窗口 3.0 秒,噪声阈值 40%,基线窗口 2 分钟,峰分割值 2,峰面积大于 2000 判定为有效积分峰。
转录组测序
使用植物通用 RNA 提取试剂盒提取样品总 RNA,借助 RNA Nano 6000 检测试剂盒与 2100 生物分析仪鉴定 RNA 完整性。取 0.5 μg 合格 RNA 反转录合成 cDNA,去除残留 RNA 后完成末端修复、加 A 尾、连接测序接头,经纯化扩增构建转录组文库,利用 Illumina NovaSeq 6000 平台上机测序。原始测序数据已上传至国家基因组科学数据中心基因组序列归档库,登录编号:CRA008967。采用 HISAT2 与 Samtools 软件将转录组测序读段比对至人参参考基因组完成序列比对分析。
原生质体制备与单细胞转录组测序
取 5 g 试验材料切碎至 1 mm 左右,置于细胞壁酶解液(含 1.5% 纤维素酶、0.4% 离析酶、0.5 M 甘露醇、20 mM 氯化钾、10 mM 氯化钙、0.1% 牛血清白蛋白)中,50~60 rpm 低速振荡酶解 2.5 h 释放原生质体。酶解完成后经 40 μm 细胞筛过滤去除细胞团,120 g 室温离心 5 min,8% 甘露醇溶液洗涤 3 次,弃上清后重悬原生质体。经台盼蓝染色鉴定细胞活性,筛选活性≥80% 的原生质体,采用高通量单细胞转录组建库试剂盒构建文库。
文库经 2100 生物分析仪完成质量检测后,使用 Illumina Hiseq2000 平台开展双端测序,单细胞转录组原始数据上传至基因组序列归档库,登录编号:CRA026200。
利用 fastp 软件完成测序数据质控与低质量序列过滤,通过专属分析流程提取细胞条形码与 UMI 标签并完成校正,比对至人参参考基因组后注释基因表达信息,最终生成基因 - 细胞表达矩阵。
运用 Seurat 软件开展单细胞下游分析,合并所有样本数据,筛选表达基因数≥100 的有效细胞,完成标准化、数据均一化与降维分析;采用 CCA 整合算法整合胚性与非胚性愈伤组织单细胞数据,选取前 30 个主成分构建细胞邻近网络,分辨率设为 0.5 完成细胞分群;借助 miloR 软件完成两类材料细胞类群丰度差异分析。
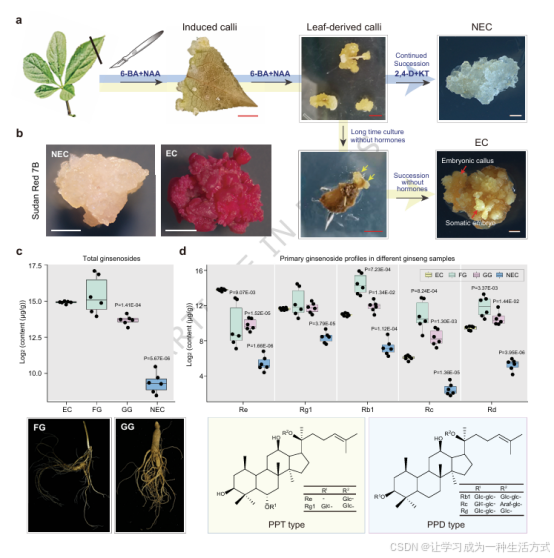
图 1 人参胚性愈伤组织(EC)具备人参皂苷合成能力
a 人参胚性愈伤组织(EC)与非胚性愈伤组织(NEC)诱导体系示意图。以人参叶片为外植体诱导获得初代愈伤组织,可进一步分化形成非胚性愈伤组织与胚性愈伤组织;其中胚性愈伤组织可继续分化产生体细胞胚与再生芽,再生能力显著优于非胚性愈伤组织。红色比例尺 = 5 mm,白色比例尺 = 1 mm。 b 人参胚性愈伤组织与非胚性愈伤组织的苏丹红 7B 染色结果。比例尺 = 0.5 cm。 c 林下参、园参、胚性愈伤组织、非胚性愈伤组织的人参皂苷总含量定量分析(n=6 生物学重复)。 d 四类材料中主要人参皂苷单体(Rg1、Re、Rb1、Rc、Rd)含量定量分析(n=6 生物学重复)。 c、d 图统计学差异采用独立双尾 t 检验;箱线图展示中位数(中线)、25%--75% 分位数(箱体)、最大最小值(须线)。原始数据已附源数据文件。
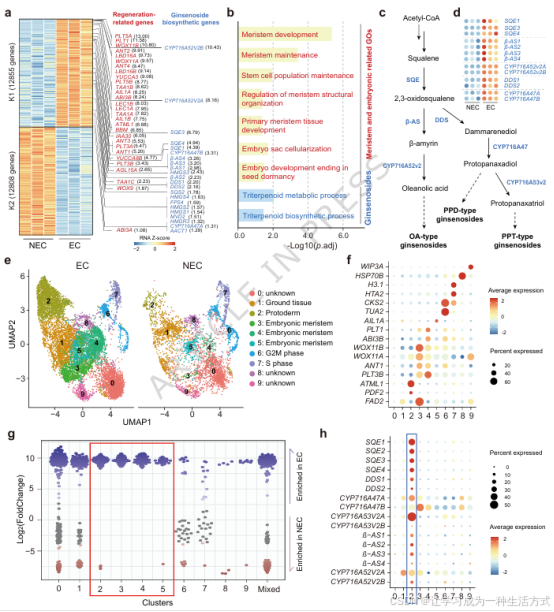
图 2 胚胎原表皮是人参皂苷合成的特异细胞来源
a 转录组鉴定的胚性 / 非胚性愈伤组织差异基因热图,基于 K-means 聚类分组。颜色代表行标准化 Z 值;标注代表性富集基因,红色为再生相关基因、蓝色为人参皂苷合成基因,并展示对应 log2 倍数变化。 b 胚性愈伤组织富集基因的 GO 功能富集分析,聚焦干细胞命运建成与人参皂苷代谢通路;显著性以校正后p 值的对数值表示(校正p <0.05)。采用超几何检验与 Benjamini--Hochberg 多重检验校正控制假阳性率。 c 人参皂苷生物合成通路示意图。 d 转录组揭示的皂苷合成关键基因(SQE 、DDS 、β-AS 、CYP716A47 、CYP716A52v2 、CYP716A53v2 )在两类愈伤组织中的表达热图。 e 单细胞转录组 UMAP 分群结果:2 组生物学重复的胚性愈伤细胞(3781、6813 个细胞)与非胚性愈伤细胞(2442、1542 个细胞)分群图谱。每个点代表单个细胞,不同颜色对应不同细胞类群,细胞类型由标志基因功能注释确定。 f 各细胞类群特异性标志基因鉴定。 g 细胞邻域丰度变化蜂群图,展示从非胚性愈伤到胚性愈伤的细胞组成重塑规律。每个点代表一组基因表达模式相近的细胞邻域(100--200 个细胞);Y 轴为胚性细胞富集的 log2 倍数变化,正值代表胚性细胞富集、负值代表耗减。颜色代表富集显著性:灰色无显著差异、蓝色显著富集、红色显著减少。 h 单细胞转录组揭示的皂苷合成基因(CYP716A52v2 、CYP716A53v2 、SQE )在 10 类细胞群中的表达模式。点大小代表表达细胞占比,颜色代表基因平均表达量。
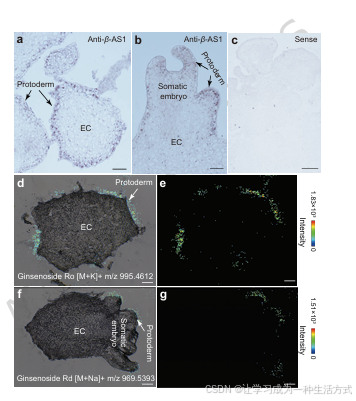
图 3 人参胚性愈伤组织原表皮特异性合成人参皂苷
a 未成熟体细胞胚中 β-AS1 反义探针原位杂交结果。 b 成熟体细胞胚中 β-AS1 反义探针原位杂交结果。 c 胚性愈伤组织中 β-AS1 正义探针原位杂交对照。 d、e AP-SMALDI 质谱成像检测皂苷 Ro 的空间分布:明场成像与质谱信号叠加图(d)、单独质谱信号图(e)。 f、g 皂苷 Rd 的质谱成像空间分布:明场成像与质谱信号叠加图(f)、单独质谱信号图(g)。 所有子图比例尺 = 200 μm。
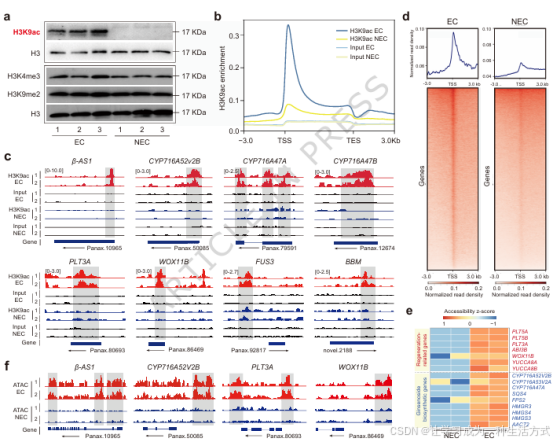
图 4 染色质开放状态决定胚性愈伤组织的皂苷合成能力
a 胚性、非胚性愈伤组织组蛋白修饰蛋白免疫印迹检测,包括 H3K9ac、H3K4me3、H3K9me2 及组蛋白 H3 内参。 b 全基因组 H3K9ac 修饰分布全景图,对比胚性与非胚性愈伤的富集信号(IP 富集组与 Input 对照组)。 c 8 个关键功能基因的 H3K9ac ChIP-seq 富集峰轨迹,包括皂苷合成基因(β-AS1 、CYP716A52v2B 、CYP716A47A 、CYP716A47B )与体细胞胚胎发生基因(PLT3A 、WOX11B 、FUS3 、BBM ),黑色标注基因区域、阴影标注差异富集峰。 d ATAC-seq 展示两类愈伤组织的全基因组染色质可及性差异,曲线为转录起始位点上下游的染色质开放信号分布,数值越高代表染色质开放程度越高。 e 再生及皂苷合成相关基因的差异开放区域热图,颜色代表可及性 Z 值;右侧标注代表性基因(红色:再生基因;蓝色:皂苷合成基因)。 f 代表性基因(β-AS1 、CYP716A52v2B 、PLT3A 、WOX11B )的 ATAC-seq 信号轨迹对比。
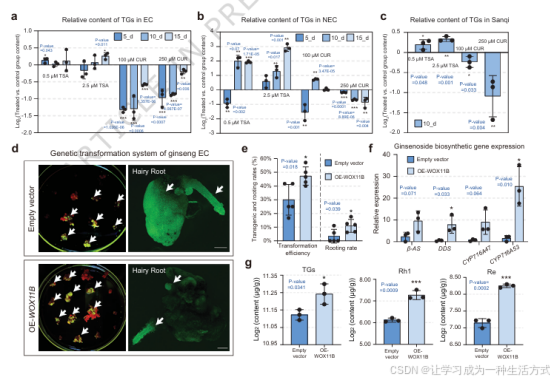
图 5 TSA 处理与 WOX11B 过表达可强化人参细胞工厂的皂苷合成效率
a 不同浓度 TSA(0.5、2.5 μM)与姜黄素 CUR(100、200 μM)分别处理 5、10、15 d 后,胚性愈伤组织总皂苷相对含量动态变化。相对含量以 log2(处理组 / 对照组)计算,对照组为 DMSO 处理。 b 相同处理条件下非胚性愈伤组织总皂苷含量动态变化。 c TSA 与 CUR 处理 10 d 后三七总皂苷相对含量变化,验证调控机制保守性。 d 空载对照与WOX11B 过表达胚性愈伤组织的 GFP 荧光转化效率检测。比例尺 = 100 μm。 e 发根农杆菌 C58C1 侵染后,两类载体材料的转化效率与生根率统计,柱上标注阳性数量(红色)与总数量(蓝色)。 f qPCR 检测空载与WOX11B 过表达株系中皂苷合成关键基因(β-AS 、CYP716A47 、CYP716A53 、DDS )的相对表达量。 g 空载与WOX11B 过表达愈伤组织中总皂苷、Rh1、Re 的相对积累量。相对含量以 log2 标准化。 a、b、c、f、g 实验设置 3 组生物学重复,e 设置 5 组生物学重复;数据为平均值 ± 标准差,采用独立双尾 t 检验统计差异。原始数据已附源数据文件。
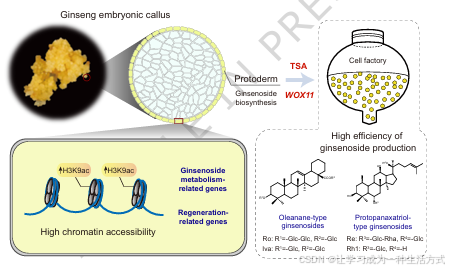
图 6 创新分子工具的开发:助力人参细胞工厂构建,实现人参皂苷高产并提升植株再生能力
高染色质开放程度与活性染色质特征,能够促进人参胚性愈伤组织中人参皂苷代谢相关基因与再生相关基因的表达。愈伤组织外层的原表皮细胞具备成为高产人参皂苷细胞工厂 的潜力。利用WOX11这类基因调控手段与 ** 曲古抑菌素 A(TSA)** 这类表观遗传调控手段,可有效提升人参皂苷的生物合成效率。