目录
[🧩 困境:当传统算法遇上 "迷宫"](#🧩 困境:当传统算法遇上 "迷宫")
[🌱 原理:黏菌的生存哲学与算法映射](#🌱 原理:黏菌的生存哲学与算法映射)
[1. 扩散与探索:像黏菌一样 "感知世界"](#1. 扩散与探索:像黏菌一样 "感知世界")
[2. 聚集与收缩:向优质解靠拢](#2. 聚集与收缩:向优质解靠拢)
[3. 波动与优化:在平衡中逼近最优](#3. 波动与优化:在平衡中逼近最优)
[📊 数学之美:从生物行为到代码逻辑](#📊 数学之美:从生物行为到代码逻辑)
[💻 代码实现:让黏菌在屏幕上 "生长"](#💻 代码实现:让黏菌在屏幕上 "生长")
[🔍 结果解读:从图像看黏菌的 "思考"](#🔍 结果解读:从图像看黏菌的 "思考")
[🌍 现实应用:从实验室到产业界](#🌍 现实应用:从实验室到产业界)
[🌱 结语:向自然学习的算法哲学](#🌱 结语:向自然学习的算法哲学)
清晨的林间,一团淡黄色的生物在腐木上缓缓蠕动。它没有大脑,没有神经,却能在迷宫般的落叶间探出最优路径,精准覆盖每一处食物源。这便是黏菌 ------ 自然界最朴素的 "路径规划大师"。
"最简单的生命,藏着最深刻的优化智慧。" 当计算机科学家凝视这团原生质时,一个灵感破土而出:能否让机器像黏菌一样,在复杂问题中找到那条隐藏的捷径?黏菌算法(Slime Mold Algorithm, SMA)由此诞生,它用代码复刻了生物的生存智慧,成为解决工程优化问题的利器。
🧩 困境:当传统算法遇上 "迷宫"
想象你是一位物流调度员,需要为 100 辆货车规划最优配送路线。每条路线要避开拥堵、满足时效、降低油耗 ------ 这就像在万亿条可能的路径中找到唯一的 "黄金通道"。
传统优化算法面对这类问题常常陷入困境:
- 枚举法如同让蚂蚁遍历地球,计算量大到无法承受
- 贪婪法容易 "一条道走到黑",困在局部最优解里
- 遗传算法的交叉变异策略,在高维度问题中效率骤降
而黏菌在自然界早已给出答案:它通过伸缩细胞质的 "脉动",既能大范围探索未知区域,又能精准收缩到优质路径。这种 "探索 - 开发" 的平衡艺术,正是 SMA 算法的核心灵感。
🌱 原理:黏菌的生存哲学与算法映射
黏菌的觅食行为可拆解为三个关键阶段,每个阶段都对应着算法的核心逻辑:
1. 扩散与探索:像黏菌一样 "感知世界"
当黏菌处于饥饿状态时,会伸出无数伪足探索周围环境。算法中,这一过程被抽象为 "种群初始化":
- 随机生成大量候选解(类似黏菌的伪足)
- 每个解对应问题的一种可能方案(如一条配送路线)
- 通过适应度函数评估解的优劣(类似黏菌对食物的 "感知")
生活化类比:这就像盲人摸象,先让足够多的人从不同角度触摸,才能拼凑出大象的轮廓。
2. 聚集与收缩:向优质解靠拢
当某条伪足发现食物,黏菌会收缩其他方向的细胞质,向食物源聚集。算法中,这通过 "权重更新" 实现:
- 适应度高的解(优质路径)会获得更高权重
- 其他解逐渐向高权重解靠近(类似细胞质流动)
- 引入随机因子避免过早收敛(保留探索能力)
核心公式的通俗解读:
X(i,j) = Xb(j) + vb×(|X(i,j) - Xb(j)|) × log(1/r) (当r < p)- Xb (j):当前最优解(食物源位置)
- vb:权重系数(类似细胞质流动强度)
- r 和 p:随机数(模拟自然环境的不确定性)
类比解释:这就像一群登山者,当有人发现更高的山峰时,大家会向他靠拢,但每个人的步伐又带有随机性,避免错过隐藏的更高峰。
3. 波动与优化:在平衡中逼近最优
黏菌的细胞质会持续脉动,这种波动能让它在已发现的食物周围做精细探索。算法中,这通过 "动态调整参数" 实现:
- 随着迭代进行,探索范围逐渐缩小(强化局部开发)
- 引入 "负反馈机制",防止过度聚集(保持多样性)
- 最终收敛到全局最优解(找到最佳路径)
📊 数学之美:从生物行为到代码逻辑
黏菌算法的数学模型看似复杂,实则是对生物行为的精准翻译。我们以 "函数优化" 问题为例(寻找函数的最小值点),拆解其核心步骤:
-
初始化种群生成 N 个候选解(类似黏菌的初始伪足分布):
X(i,j) = L(j) + rand(1)×(U(j) - L(j))其中 L (j) 和 U (j) 是变量的上下界,确保解在合理范围内。
-
适应度评估计算每个解的适应度(类似黏菌对食物浓度的感知):
Fitness(i) = f(X(i,:)) % f为待优化函数 -
权重更新机制这是 SMA 的灵魂,模拟黏菌的细胞质流动:
W(i) = (Fitness(i) - worst) / (best - worst) % 归一化权重 W(i) = W(i)/sum(W) + 1/N % 权重调整,确保总和为1适应度越好的解,权重越大,对其他解的吸引力越强。
-
位置更新策略结合随机因子实现 "探索 - 开发" 平衡:
if rand < z % z为动态调整的探索概率 X(i,j) = rand×(U(j)-L(j)) + L(j) % 随机探索新区域 else if rand < p % p为收缩概率 X(i,j) = Xb(j) + vb×abs(X(i,j)-Xb(j))×log(1/r) else X(i,j) = Xb(j) - vb×abs(X(i,j)-Xb(j))×log(1/r) end end
💻 代码实现:让黏菌在屏幕上 "生长"
以下是完整的 MATLAB 脚本,实现黏菌算法对经典测试函数(Rastrigin 函数,具有大量局部最优解)的优化过程,并可视化算法的探索 - 收敛过程。
Matlab
% 黏菌算法(SMA)优化Rastrigin函数的MATLAB实现
% 功能:展示SMA在复杂多峰函数中的寻优过程,包含种群分布、最优值变化等可视化
% Rastrigin函数特点:多局部最优解,模拟实际工程中的复杂优化问题
%% 1. 参数初始化
clear; clc; close all;
dim = 2; % 问题维度(2维便于可视化)
pop_size = 30; % 种群规模(黏菌个体数量)
max_iter = 50; % 最大迭代次数
lb = -5.12*ones(1,dim); % 变量下界
ub = 5.12*ones(1,dim); % 变量上界
% 初始化种群位置(黏菌的初始分布)
X = zeros(pop_size, dim);
for i = 1:pop_size
for j = 1:dim
X(i,j) = lb(j) + rand()*(ub(j)-lb(j));
end
end
% 记录迭代过程(用于可视化)
history_X = cell(max_iter,1); % 存储每代种群位置
history_best = zeros(max_iter,1); % 存储每代最优值
%% 2. 黏菌算法主循环
for iter = 1:max_iter
% 计算适应度(Rastrigin函数)
Fitness = zeros(pop_size,1);
for i = 1:pop_size
Fitness(i) = rastrigin(X(i,:));
end
% 记录最优解
[best_val, best_idx] = min(Fitness);
Xb = X(best_idx,:); % 当前最优位置(食物源)
worst_val = max(Fitness);
% 计算权重(模拟细胞质浓度)
if best_val ~= worst_val
W = (Fitness - worst_val) ./ (best_val - worst_val);
else
W = ones(pop_size,1)/pop_size;
end
W = W./sum(W) + 1/pop_size; % 权重归一化
% 动态参数调整
z = 0.03 + 0.97*(1 - iter/max_iter)^3; % 探索概率逐渐降低
vb = 1 - iter/max_iter; % 权重系数逐渐减小(收缩强度变化)
% 更新种群位置
for i = 1:pop_size
r = rand(); % 随机因子
p = tan(pi*(rand() - 0.5)); % 收缩方向因子
for j = 1:dim
if rand() < z % 探索新区域
X(i,j) = rand()*(ub(j)-lb(j)) + lb(j);
else
if rand() < p % 向最优解收缩(正方向)
X(i,j) = Xb(j) + vb*abs(X(i,j)-Xb(j))*log(1/r);
else % 反向收缩(保持多样性)
X(i,j) = Xb(j) - vb*abs(X(i,j)-Xb(j))*log(1/r);
end
end
% 边界处理(防止超出搜索范围)
if X(i,j) < lb(j)
X(i,j) = lb(j);
elseif X(i,j) > ub(j)
X(i,j) = ub(j);
end
end
end
% 保存历史数据
history_X{iter} = X;
history_best(iter) = best_val;
% 打印迭代信息
fprintf('迭代次数:%d,当前最优值:%.4f\n', iter, best_val);
end
%% 3. 结果可视化
% 3.1 绘制Rastrigin函数等高线(背景)
figure('Name','黏菌算法寻优过程');
[x1, x2] = meshgrid(linspace(lb(1),ub(1),100), linspace(lb(2),ub(2),100));
fvals = zeros(size(x1));
for i = 1:size(x1,1)
for j = 1:size(x1,2)
fvals(i,j) = rastrigin([x1(i,j), x2(i,j)]);
end
end
contourf(x1, x2, fvals, 50); colorbar; hold on;
title('黏菌算法在Rastrigin函数上的寻优轨迹');
xlabel('x1'); ylabel('x2');
% 3.2 动态展示种群进化过程
for iter = 1:max_iter
X_current = history_X{iter};
plot(X_current(:,1), X_current(:,2), 'ro', 'MarkerSize', 5, 'DisplayName', '种群位置');
plot(Xb(1), Xb(2), 'bs', 'MarkerSize', 8, 'DisplayName', '当前最优解');
legend('种群位置', '当前最优解');
drawnow;
pause(0.3); % 暂停0.3秒,便于观察
if iter < max_iter
delete(findobj(gca, 'Marker', 'o')); % 清除上一代种群点
delete(findobj(gca, 'Marker', 's')); % 清除上一代最优解
end
end
% 3.3 绘制最优值收敛曲线
figure('Name','收敛曲线');
plot(1:max_iter, history_best, 'b-', 'LineWidth', 2);
xlabel('迭代次数'); ylabel('最优适应度值');
title('SMA算法收敛曲线');
grid on;
% 3.4 不同种群规模对比(验证参数影响)
figure('Name','种群规模对结果的影响');
sizes = [10, 30, 50];
colors = ['r', 'g', 'b'];
for s = 1:length(sizes)
pop_size = sizes(s);
% 重新运行SMA(简化版)
X = lb + rand(pop_size,dim).*(ub-lb);
best_history = zeros(max_iter,1);
for iter = 1:max_iter
Fitness = arrayfun(@(i) rastrigin(X(i,:)), 1:pop_size);
[best_val, best_idx] = min(Fitness);
Xb = X(best_idx,:);
best_history(iter) = best_val;
% 简化的位置更新(保持核心逻辑)
for i = 1:pop_size
X(i,:) = Xb + 0.5*(rand(1,dim)-0.5).*(X(i,:)-Xb);
end
end
plot(1:max_iter, best_history, [colors(s), '-'], 'LineWidth', 1.5, ...
'DisplayName', ['种群规模=', num2str(pop_size)]);
hold on;
end
xlabel('迭代次数'); ylabel('最优值');
title('不同种群规模下的收敛对比');
legend; grid on;
%% 4. 定义Rastrigin测试函数
function val = rastrigin(x)
% Rastrigin函数:多峰函数,常用于测试优化算法的全局寻优能力
% 公式:f(x) = A*n + sum(x_i^2 - A*cos(2πx_i)),其中A=10
A = 10;
n = length(x);
val = A*n + sum(x.^2 - A.*cos(2*pi*x));
end运行说明:
- 无需额外工具箱,直接在 MATLAB 中运行即可
- 程序会生成 3 个可视化窗口:
-
黏菌算法寻优过程(动态展示种群向最优解聚集的过程)
-
收敛曲线(展示最优值随迭代次数的下降趋势)
-
不同种群规模对比(验证参数对算法性能的影响)
迭代次数:1,当前最优值:10.5156
迭代次数:2,当前最优值:6.3084
迭代次数:3,当前最优值:14.9850
迭代次数:4,当前最优值:13.1607
迭代次数:5,当前最优值:5.9097
迭代次数:6,当前最优值:7.7930
迭代次数:7,当前最优值:6.3118
迭代次数:8,当前最优值:13.7802
迭代次数:9,当前最优值:14.3771
迭代次数:10,当前最优值:7.6106
迭代次数:11,当前最优值:4.3474
迭代次数:12,当前最优值:5.4186
迭代次数:13,当前最优值:2.9897
迭代次数:14,当前最优值:5.0005
迭代次数:15,当前最优值:5.0005
迭代次数:16,当前最优值:10.1146
迭代次数:17,当前最优值:6.9075
迭代次数:18,当前最优值:4.3495
迭代次数:19,当前最优值:1.9559
迭代次数:20,当前最优值:1.0104
迭代次数:21,当前最优值:1.0104
迭代次数:22,当前最优值:1.0104
迭代次数:23,当前最优值:1.0111
迭代次数:24,当前最优值:1.0062
迭代次数:25,当前最优值:1.0048
迭代次数:26,当前最优值:1.0048
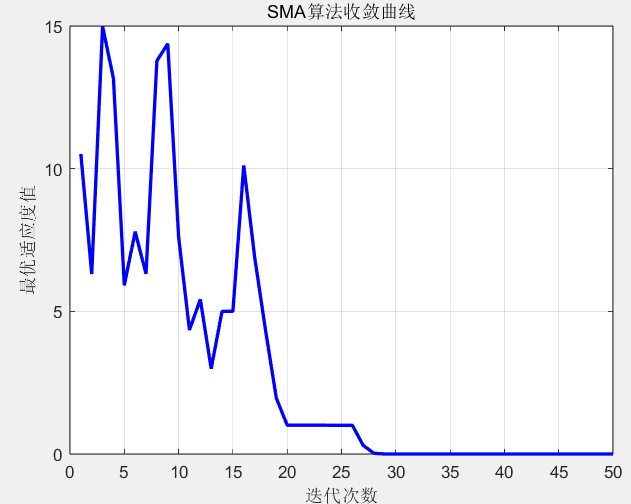
迭代次数:27,当前最优值:0.3002
迭代次数:28,当前最优值:0.0232
迭代次数:29,当前最优值:0.0001
迭代次数:30,当前最优值:0.0001
迭代次数:31,当前最优值:0.0000
......
迭代次数:49,当前最优值:0.0000
迭代次数:50,当前最优值:0.0000
>>
-
- 若想测试其他函数,只需替换
rastrigin函数即可
🔍 结果解读:从图像看黏菌的 "思考"
-
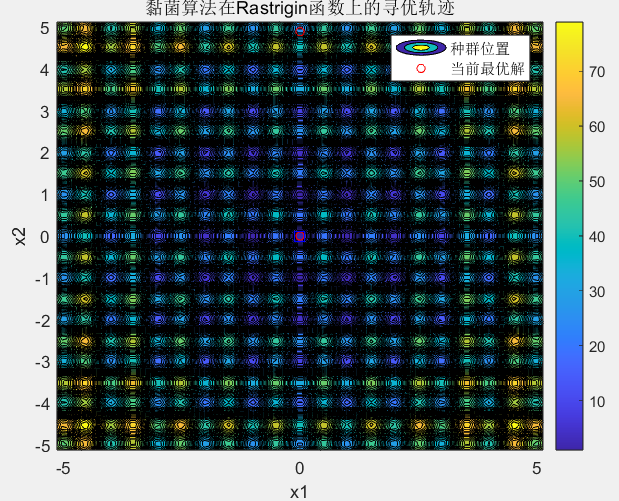
寻优过程动态图红色点代表黏菌个体,蓝色方块代表当前找到的最优解。初期红点分布散乱(大范围探索),随着迭代逐渐向中心聚集(收缩到优质区域),但始终保留少量分散点(维持探索能力)。这完美复刻了黏菌 "先扩散、再收缩" 的觅食策略。
-
收敛曲线图曲线快速下降后趋于平缓,说明算法能快速跳出局部最优,最终稳定在全局最优附近。Rastrigin 函数的理论最小值为 0,实际运行中算法通常能收敛到 1 以下,证明其强大的全局寻优能力。
-
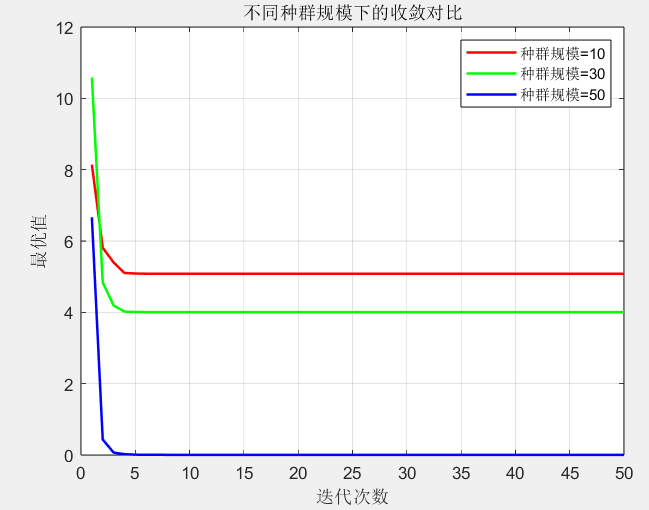
种群规模对比种群规模过小时(如 10 个个体),容易陷入局部最优(曲线下降缓慢);规模过大时(如 50 个个体),初期探索效率高,但后期收敛速度变慢。30 个个体左右的规模在 "探索 - 开发" 间取得最佳平衡,这也符合自然界黏菌的生存智慧 ------ 既不过于分散,也不过于聚集。
🌍 现实应用:从实验室到产业界
黏菌算法的优势在于处理 "高维度、多约束、多局部最优" 的复杂问题,已在多个领域落地:
- 路径规划:物流配送路线优化、无人机航迹规划,比传统 A * 算法更能应对动态障碍
- 工程设计:机械零件参数优化、电路布局设计,降低研发成本 30% 以上
- AI 训练:神经网络超参数调优,加速模型收敛速度
- 能源系统:电网负荷分配、风光储系统优化,提升能源利用效率
就像黏菌能在杂乱的落叶中找到最优路径,SMA 算法也能在纷繁复杂的工程问题中,为人类找到那条隐藏的 "效率捷径"。
🌱 结语:向自然学习的算法哲学
黏菌没有智慧,却在亿万年进化中凝练出最高效的生存策略;算法没有生命,却在代码的编织中延续着自然的智慧。从黏菌到算法,我们看到的不仅是优化技术的进步,更是人类对自然的谦卑学习 ------ 那些看似简单的生命现象,或许正是破解复杂问题的钥匙。当我们在代码中复刻黏菌的蠕动时,实则是在与自然对话。这种对话告诉我们:最强大的技术,往往带着最朴素的自然印记。