本笔记基于杨荣武教授主编的《生物化学》第四版,配图也源于教材~
一、脂肪酸的分解
脂肪酸的分解是以氧化的形式进行的,根据氧化的碳原子的位置可以分为α氧化、β氧化和omega氧化。其中β氧化是最主要的形式,α氧化是处理β碳出现甲基相连的情况,我们对omega氧化不做介绍(简单知道是从最后一个碳原子开始氧化,而且这种氧化是不需要活化脂肪酸的即可)
脂肪酸的β氧化
书上在介绍脂肪酸的β氧化的时候有一个引入,是如何发现β氧化是以两碳形式逐个进行的,这个当时是利用了苯环作为脂肪酸氧化的停止点,那么发现在奇数脂肪酸是生成苯甲酸衍生物,而偶数脂肪酸是生成苯乙酸衍生物(两点需要注意,一是这里的偶数和奇数是计算了羧基的碳的,二是如果不连接苯环正常氧化直链脂肪酸的话,那么偶数脂肪酸会全部转变为乙酰CoA,奇数脂肪酸会生成一个丙酰CoA,其他的碳生成乙酰CoA)
另外一个非常重要的事实的,脂肪酸的β氧化是主要发生在线粒体基质当中(后面会有补充)
β氧化的历程
其实β氧化的历程有点类似柠檬酸脱氢转变为顺乌头酸然后水合变为异柠檬酸最后异柠檬酸脱羧,这里β氧化也基本上遵循着活化(转变为脂酰辅酶A),脱氢,水合,裂解的过程
(1)脂肪酸的活化
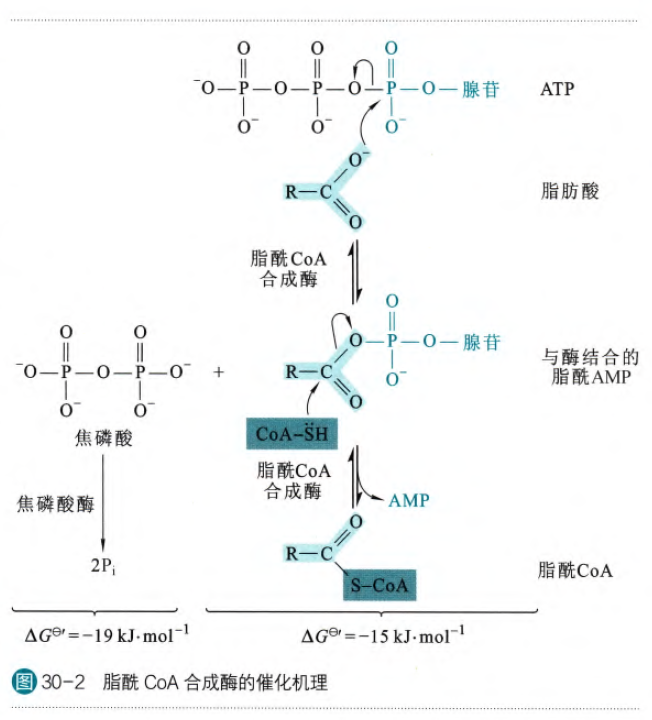
脂肪酸在脂酰辅酶A合成酶的作用下消耗一个ATP生成脂酰辅酶A。这里可以理解为为脂肪酸首先加上了AMP,然后生成了一个焦磷酸(焦磷酸极易水解生成两个磷酸基团,这一步会释放较多自由能),然后辅酶A替代AMP生成了脂酰辅酶A。
催化这步反应的脂酰辅酶A合成酶也称为硫激酶,有几种不同的类型,有活化长链脂肪酸的分布在线粒体外膜和内质网膜上,有处理中短链脂肪酸的分布在线粒体内膜的外侧。
这个反应特别需要注意的虽然只有一个ATP参与反应,但是这个反应实际上消耗了2个ATP,因为实际上是水解了两个高能磷酸键
(2)脂酰辅酶A的转运
为什么要转运呢?因为细胞当中储存脂肪都是在细胞质当中的,但是脂肪酸最后分解产生的乙酰CoA是需要在线粒体基质当中运用的,那么最优的方式自然是把原料运输到线粒体基质当中(从脂肪CoA合成酶的细胞定位也可以知道,我们必须要经历这一步)
但是脂酰CoA不可以直接通过线粒体内膜,主要的限制因素在CoA上,CoA是一个强极性的基团,这里需要借助三个重要的蛋白分别是肉碱-软脂酰转移酶I(CPTI)和肉碱-软脂酰转移酶II(CPTII)和脂酰肉碱转位酶(CACT)
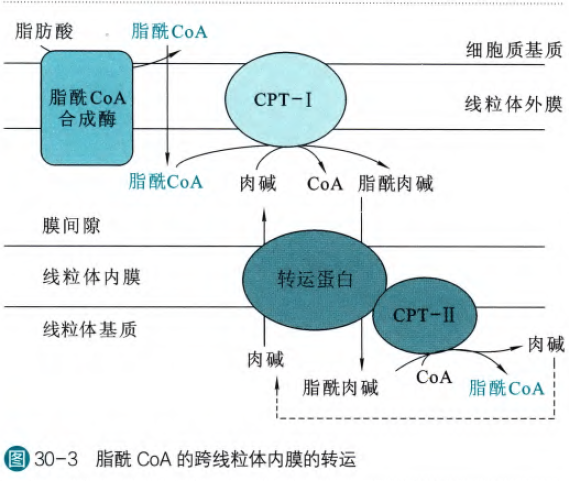
若脂酰CoA合成酶分布在线粒体外膜上(如果不在也可以脂酰CoA通过转运至外膜),那么合成的脂酰CoA可以透过外膜进入线粒体膜间隙当中,在CPT-I的作用下,肉碱替代CoA形成脂酰肉碱,然后脂酰肉碱可以通过图上标出的转运蛋白(即脂酰肉碱转位酶)运输到线粒体基质当中,然后在CPT-II的作用下肉碱被CoA替代重新生成脂酰CoA。但是我们会发现这样一个问题,那不是在线粒体基质当中就会囤积肉碱吗?其实不然,在脂酰肉碱转位酶转运的时候,实则是将肉碱和脂酰肉碱1:1同时反向转运,从而维持这个转运体系的运转。
所以肉碱在脂肪酸的氧化上起到了非常重要的的作用,我们可以通过食物获得充足的肉碱,我们也可以自己合成肉碱,所以一般不会因为缺乏肉碱致病。但是虽然肉碱可以促进脂肪酸的氧化,但是也没有多额外摄入的必要。
(3)脱氢
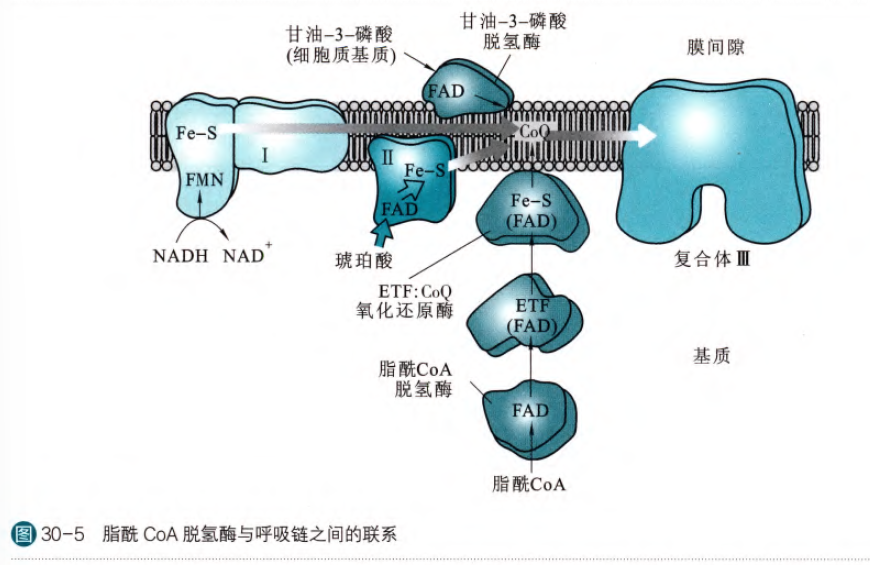
这一步是由脂酰CoA脱氢酶催化的,此脱氢酶依赖FAD,FADH2将电子通过电子传递黄素蛋白,传递给铁硫蛋白然后最后传递给辅酶Q,然后从复合体III进入呼吸链。所以这里的FADH2不通过复合体II进入呼吸链,而是直接从复合体III进入,所以这一步会生成1.5ATP
这一步的脂酰CoA脱氢酶存在三种,分别作用于短链(2~5),中链(6~12)和长链(13~21),其中处理短链和中链的是定位在线粒体基质中,处理长链的是定位在线粒体内膜上。
荔枝当中含有一种物质称为降糖氨酸A,这种分子可以强力抑制处理中链和短链的脂酰CoA脱氢酶活性,从而抑制脂肪酸的分解。那么脂肪就会积累而无法利用,这时候细胞会通过加快糖的吸收分解来代偿,这样做会有助于血糖的降低。但是因为降糖氨酸A同时抑制了糖异生的能量来源(糖异生不可能用着分解糖产生的能量然后合成糖吧~),所以肝细胞就不能再通过糖异生向外输送葡萄糖,这样进一步导致血糖的降低。所以荔枝最好不要空腹食用,因为空腹时本身肝糖原储存不多,这样可能会过度消耗肝糖原,最后导致低血糖。
(4)加水
我们在上一步生成了一个反式的双键,这一步通过水合反应,在β碳原子上加上羟基,同时因为此时β碳原子是手性碳,所以有必要强调我们生成的都是L型的β-羟酰CoA。这一步反应由烯酰CoA水合酶催化
(5)再脱氢
这一步将上一步加上的羟基进行进一步氧化,生成羰基,这一步反应由羟酰CoA脱氢酶催化,此脱氢酶依赖辅酶I,故生成一分子NADH,因为此反应发生在线粒体基质,所以生成的NADH中的高能电子可以直接通过复合体I进入呼吸链
(6)硫解
这一步反应由硫解酶催化。首先酶活中心的半胱氨酸的巯基进攻β-碳,然后释放出乙酰CoA,然后新的CoA再次进攻β-碳,替代巯基,最后生成少两个碳的新的脂酰CoA
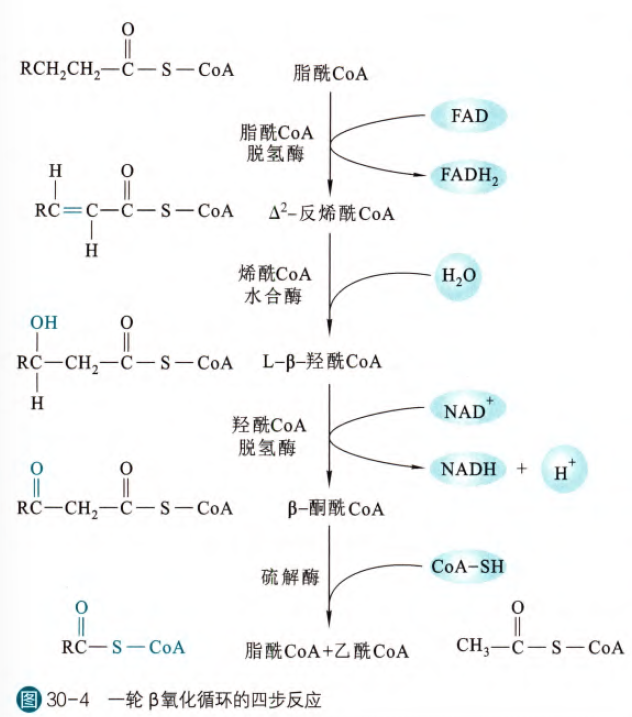
这是脂肪酸经过活化和转运之后不断发生的一轮β氧化循环的过程,那么在这个过程当中涉及的能量:一分子的FADH2和一分子的NADH,通过呼吸链可以生成4个ATP,那么以软脂酸为例(16碳),就可以生成28个ATP,然后一共生成8个乙酰CoA,故通过三羧酸循环和呼吸链一共生成80个ATP,最后因为最初在活化脂肪酸的时候消耗了两个ATP,所以一分子软脂酸可以生成106个ATP。
书上还问了这样一个问题,一分子乙酸可以生成多少ATP?其实基本原理是一样的,首先乙酸也需要活化生成乙酰辅酶A,这一步仍然需要2个ATP,接着再转运到线粒体基质当中,通过TCA和氧化呼吸链得到10个ATP,所以总共可以生成8个ATP。那么接着再问,一分子乙醇可以生成多少ATP,这里可以把问题转化为将乙醇氧化为乙酸可以产生多少ATP,答案如果用NADH表示很简单即为2分子,但是因为乙醇的氧化是在细胞质基质当中进行的,所以NADH的电子转运可能会走MAP途径,也可GAP途径,所以最后可以生成13/12/11个ATP
其实通过上面的反应我们可以发现实际上,我们所做的两步氧化的对象都是β碳,所以我们说这事β氧化,那么实际上我们氧化得到的β碳是作为新的CoA连接的接头,而不是直接生成乙酰CoA。那么生成的乙酰CoA在三羧酸循环里面释放的能量主要都是来自于未被氧化的α碳,所以简单理解为β碳在进入三羧酸循环前利用,α碳在三羧酸循环里面氧化利用。
那么如果我们用一个整体的视角去看整个脂肪酸分解的反应,实际上就是脂肪酸与氧气发生氧化还原反应释放的能量,脂肪酸原本的还原C的电子全部转到电子传递链上释放能量,而所有的碳都在TCA当中经过脱羧反应以CO2形式释放(以软脂酸为例)。另外,其实所有的氢也最终和氧气结合形成了水,实际上骆驼长途跋涉利用的水就是脂肪酸代谢产生的水。
那么当我们了解到β氧化的过程之后,我们处理以下几个问题:
问题:
(1)奇数脂肪酸的β氧化如何进行
我们之前一直是以软脂酸为例,但是如果我们分解的是奇数脂肪酸呢?实际上在前面的反应当中都是正常进行活化和β-氧化的,只是在最后的β碳上我们发现实际上连接了最末端的甲基,这种情况其实就已经到达反应的终点,最后生成丙酰CoA。但是其实我们在糖异生当中提到过丙酰CoA,这是糖异生的原料,那丙酰CoA经过了什么反应加入到糖异生中呢?实则是经过羧化和异构反应,最后转变为琥珀酰CoA,从而既可以参加三羧酸循环,也可以转变到草酰乙酸之后离开线粒体,参与糖异生途径。下面是丙酰CoA转变为琥珀酰CoA的历程。简单来说就是先羧化(催化这一步反应的酶需要生物素作为辅基,同时也需要消耗一分子的ATP,类似于糖异生途径中丙酮酸的羧化),然后差向异构,之后再将羧基变位,得到琥珀酰辅酶A。
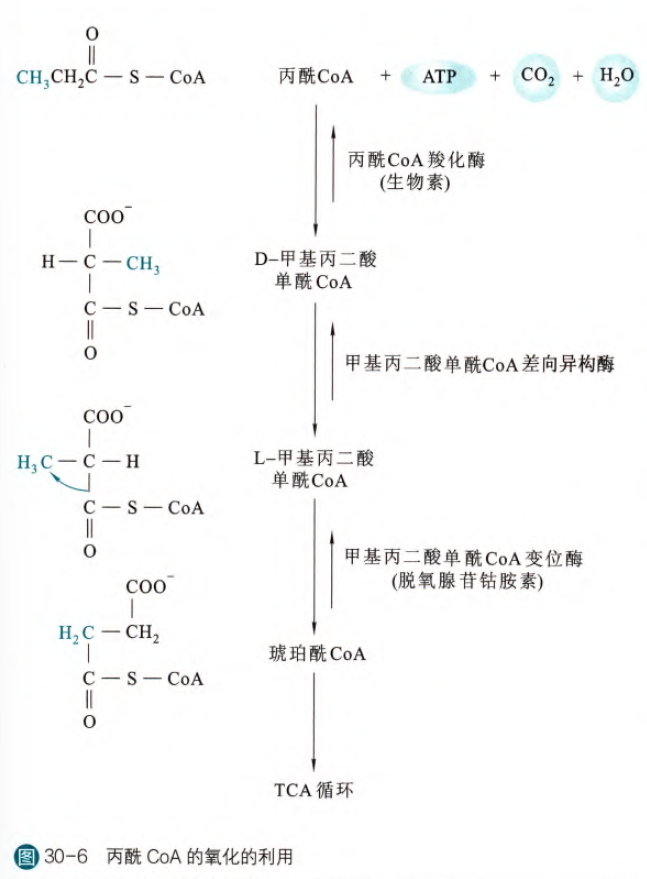
需要注意的是,催化第二步变位反应的酶需要脱氧腺苷钴胺素作为其的辅酶,这种辅酶参与催化的方式也很特别,即提供自由基来辅助催化反应。(这种辅酶衍生于维生素B12(素食主义者需要注意),所以缺乏会导致低血糖和酮症的发生)
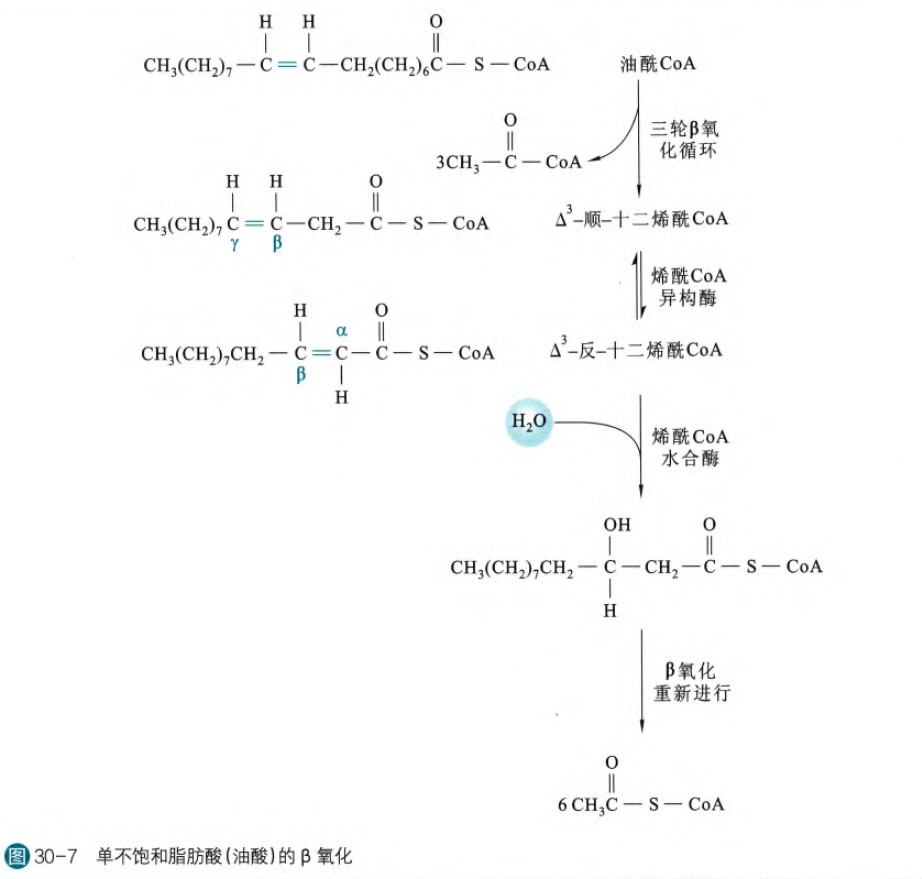
(2)不饱和脂肪酸的β氧化
如果不饱和脂肪酸的双键恰好是在反式也是正确的位点,那么实际上也就是跳过了最开始的脱氢步骤,仅仅是少了1.5个ATP,不会干扰后面的反应。但是我们摄入的不饱和脂肪酸绝大多数都是顺式(自然界的都是),所以这样首先需要烯酰CoA异构酶将其变为反式,然后再正常反应(这一步也是跳过了脱氢哈~)
那么如果遇到多个不饱和键,那么需要还原酶的作用啦
(3)过氧化物酶体和乙醛酸循环体内发生的β氧化
对于动物而言,脂肪酸β氧化不仅可以在线粒体内进行,也可以在过氧化物酶体中进行,那么对于植物来说,在动物的基础上还多了一个乙醛酸循环体。
实则在不同的细胞器进行β氧化本质差别并不是很大,只是在第一步的时候,过氧化氢酶体和乙醛酸循环体将电子传递给FADH2之后,FADH2会直接将电子传递给氧气而不会经过呼吸链所以这一步会少1.5个ATP。还有一个差别在于,因为在过氧化物酶体和乙醛酸循环体内产生的乙酰CoA和NADH都是要在线粒体当中用的,NADH即可以通过我们之前说过的苹果酸途径或者天冬氨酸途径转出细胞器,乙酰CoA可以用我们刚才介绍的肉碱转运体系运出细胞器。
线粒体一般不会处理超长链脂肪酸,这种脂肪酸一般就会交给过氧化物酶体和乙醛酸循环体处理。这种超长链脂肪酸进入过氧化物酶体不需要肉碱,实际上通过一种主动运输蛋白运入的。如果这种转运蛋白缺失可能就会引起超长链脂肪酸在体内的积累,患上脑白质肾上腺营养不良症。
但是过氧化物酶体的β氧化酶系对短链脂肪酸不起作用,所以只能交给线粒体继续进行(乙醛酸循环体可以善始善终)
脂肪酸的α氧化
什么时候会需要用到脂肪酸的α氧化呢?就是在β碳上连接了一个甲基(可能是因为异戊二烯引入的),这个时候就无法进行β氧化了,所以需要α氧化这个碳,从而去除分支,使得β氧化继续进行(但是如果α碳连上甲基是可以进行β氧化的,但是最终是生成丙酰CoA)
那么我们以植烷酸为例:
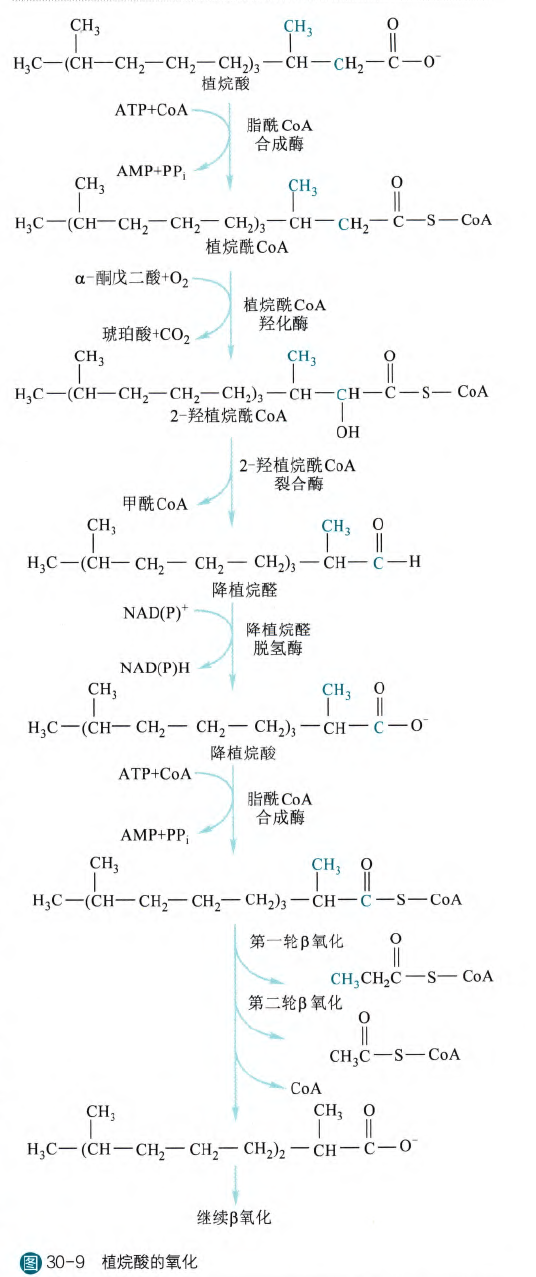
得到植烷酰CoA之后首先植烷酰羟化酶在α碳上加上一个羟基(这一步需要α-酮戊二酸和氧气的参与),然后直接在裂合酶的作用下直接分裂得到甲酰CoA和降植烷醛,之后在脱氢酶的作用下氧化生成降植烷酸,之后降植烷酸还是需要经历新一轮活化再参与反应。
所以实际上如果出现这种分支,处理起来是需要耗费一定能量的,虽然生成了NADH或者NADPH,但是需要经历多一轮的活化。
除此之外,我们可以敏锐地观察到,实际上消除的碳不是原来的α碳,而是羧基碳,让原来连接甲基的β碳经过这一次的反应变为连接甲基的α碳,这样就可以进行β氧化了(碳不变,只是位置变了)
酮体的生成和利用
酮体包括丙酮、乙酰乙酸和D-β-羟丁酸。酮体一般是在饥饿、高脂肪低糖饮食等情况下会大量合成,因为在这些情况中,血糖的降低使得需要在三羧酸循环里面抽调代谢物进行糖异生,这样会使得TCA的运转速度变慢,乙酰CoA积累,那么积累的乙酰CoA便会参与酮体的合成。酮体主要的合成场所是肝细胞的线粒体基质(毕竟这是糖异生的主要细胞),肾脏细胞和神经胶质细胞也可以合成少量酮体。
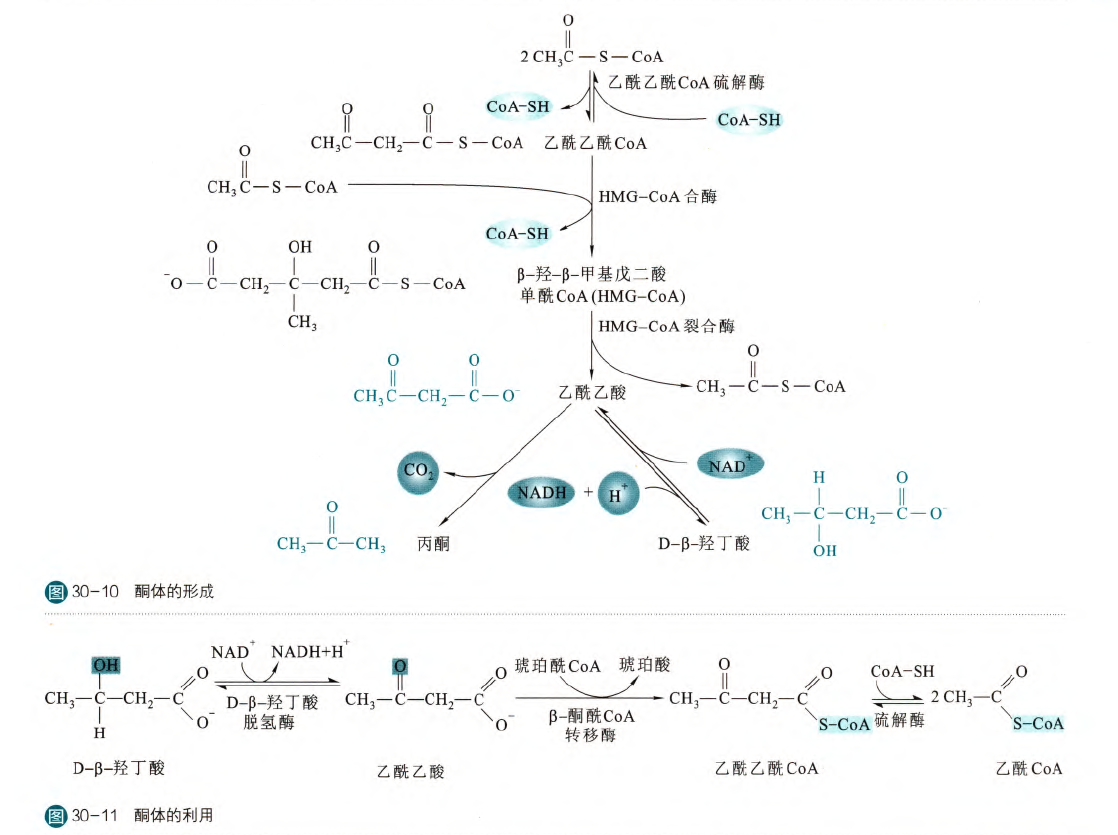
第三步生成第一个酮体------乙酰乙酸,乙酰乙酸可以自发脱羧转变为丙酮,也可以在脱氢酶的作用下还原为D-β-羟丁酸,其他上面的反应历程就不再多说,反应产生的丙酮可以通过自由扩散进入血液,然后通过呼吸呼出,所以可以通过呼出的气体来初筛糖尿病。产生的乙酰乙酸和D-β-羟丁酸需要在质膜上的单羧酸转运蛋白2(MCT2)来离开肝细胞进入血液循环,再经过MCT2被组织吸收比如脑、骨骼肌、心肌和肺。乙酰乙酸和D-β-羟丁酸可以通过β-酮酰CoA转移酶来转变为乙酰乙酰CoA,然后再通过硫解酶的逆反应产生乙酰CoA。所以在饥饿的时候,酮体就成为了脑、骨骼肌、心肌等的重要燃料之一。但是肝细胞不可以利用酮体,因为缺乏β-酮酰CoA转移酶。
虽然酮体可以作为一种备用燃料,但是因为乙酰乙酸和D-β-羟丁酸都是中强酸,所以酮体过多会产生酮症,甚至出现酸中毒,因此切不可饥饿过久或者长时间使用高脂肪低糖的食物。
(所以生成和利用酮体虽然是TCA被抑制的另一条小路,但是如果这条小路产生的酮体过多也会对机体有害)