《智能的理论》全书转至目录****
不同AGI的研究路线对比简化版:《AGI(具身智能)路线对比》,欢迎各位参与讨论、批评或建议。
神经系统分为中枢神经系统和外周神经系统。外周神经系统主要指的是的运动神经元和感觉神经元。其中,运动神经元如alpha和gamma,感觉神经元如肌梭、关节感受器和触压觉感受器等("15-4:身体感觉:本体感觉"和"15-2:身体感觉:触觉")。中枢神经系统包括大脑和脊髓,跟运动控制的相关脑区主要包括脑干、运动皮质、小脑和基底神经节。脊髓属于最低层次的中枢神经组织,处于大脑和外周神经系统之间。另外,在神经系统中还存在很多反射弧,任何反射活动(如常见的膝跳反射)都以反射弧为基础。
一.反射(张英波和夏忠梁,2019)
1.反射弧
反射是基本的神经系统活动方式。反射活动需要通过反射弧实现,它包括感受器、传入神经、反射中枢、输出神经和效应器。感受器是身体的"感觉传感器",负责上传感觉信息,比如触觉感受器等。反射中枢即中枢神经系统,它包括脊髓、小脑、中脑和大脑等,是反射的处理单元。越低级的反射中枢(脊髓)其反射越简单,越高级的反射中枢(大脑皮质)则越复杂。效应器是反射动作的实现器官,如肌肉。反射的过程首先是感受器接收感觉信息,然后通过传入神经将信号传至反射中枢,经过处理后再将处理后的信息从输出神经传至效应器完成反射。
按不同的维度反射可以有不同的分类。
2.非条件反射和条件反射
非条件反射是指与生俱来的的反射,这种反射的数量比较有限并无需大脑皮层参与;条件反射是指可以后天学习的,形式多变且复杂,需要在大脑皮层的参与下完成的。
3.M1、M2、激发性、M3反射
(1)M1反射
M1反射表示突然增加肌肉负荷使肌梭受到拉伸而引起的反射。它的感受器和效应器为同一块肌肉的梭肌和肌肉。人体内唯一的M1反射是腱反射,而大家熟悉的膝跳反射就是腱反射的一种。它有如下三个特点:
(a)它是在神经中枢中只通过一次突触传递的反射,它从感受器起最远传至脊髓。其反应时间很快,反应时间只需30-50毫秒。因为M1反射只需通过一次1突触传递,所以它也被称为单突触反射;
(b)因为M1反射只在脊髓中处理,所以它是在自动化无意识的情况下进行的。其灵活性也是最差的,一种刺激对应一种反应,只要达到它的触发条件就会触发;
(c)不受海克定理制约(指当呈现的刺激越多,个体反应的时间越长)。M1反射可以并行处理,即当不同肢体同时达到M1反射的条件,就可以同时进行多个M1反射。
(2)M2反射
M2反射需要通过多次突触传递,又称长环牵张反射。肌紧张就属于一种M2反射(在牵张反射中有介绍)。它有如下三个特点:
(a)其信号可以到达小脑、中脑和皮层组织。因此它的反应时间比M1的慢,时间需要50-80毫秒;
(b)M2反射也是可以无需意识控制的。但可以通过一定的意识对该反射进行控制,因此它的灵活性比M1反射强。比如通过意识控制,可以使肌肉放松从而可以坐下或者蹲下(调节肌紧张)。M2反射还可以通过一些自我暗示(还不算意识)调整,比如在滑雪时为了在颠簸状态下保持平稳,会在过程中随时控制膝盖作出一定的弯曲(调节肌紧张);
(c)与M1反射一样,它可以并行处理,比如在用水杯接水时,我们可以一边站着(肌紧张1),一边手握水杯接水(肌紧张2);
(d)M2反射处理的信息比M1反射更多,除了梭肌,还包括关节和姿势等信息,因此M2反射能处理更复杂的信息作出更复杂的反应。换句话说,M1反射是为了维持某组梭肌长度,而M2反射则可以对一套梭肌,关节和姿势等组织进行保持。
(3)激发性反射
这种反射是通过后天习得的,可以通过反复的体验或学习将刺激与任意的肌肉群联系。一个激发性反射的例子是,在柔道比赛中运动员抱着对手的腰,当运动员的手臂感受到对手在挣脱时,能下意识的将手抱得更紧。这些是在后天通过练习习得的。激发性反射比M2反射的反应时间长,为80-120毫秒。因为运行时间较快,所以它也可以归入到下意识处理的范畴中。但由于它具有学习性,通过后天可以学习各种各样的反射,并且它受意识的影响更大,所以它的灵活性比M1和M2都大。激发性反射发生的肌肉群能远离刺激的位置。比如左手感应而右手反应。
(4)M3反射
M3反射也称为随意反射,处理中枢是大脑,反应时间约为120-180毫秒。M3反射受意识控制,所以相比其他反射它有具最大的灵活性,能按照人的意愿进行。因为M3受意识控制,所以它只能串处理,即同一时刻能同时处理的M3反射极少,受海克定律制约。与激发性反射一样,M3反射发生的肌肉群能远离刺激的位置。M3反射的一个例子是司机通过时刻对车道线的观察(车是否在车道线间或是否压线),始终将车控制在两车道线之间。
M3反射受意识的控制,那么M3反射与随意运动有什么区别呢?反射意味着该运动受到感觉信号的反馈调控,而随意运动则可以不根据外部信息调控。所以M3反射与随意运动的区别在于是否必须根据外部的感觉信号进行调整。另外,当M3反射与激发性反射的区别在于自动化程度,激发性反射比M3反射的自动化程度更高。当随着练习逐渐变得更熟练时,M3反射可以转变为激发性反射。
4.多级反射的整合
很多反射并不是单独发生的,而是多级反射共同作用,以更好地应对外部刺激的变化(Schmidt和Wrisberg,2000)。在一个实验中,需要被试用手托住一本较重的书并要求其保持书的高度,同时利用肌电图仪(测量肌肉活动的仪器)测量手肌肉的电活动。在被试不预知的情况下,突然在被试托着的书的上面加上另外一本书。肌电图的测量情况如图1所示,最先出现的是M1反射,它对新增加的负荷作出一定调整,继而是M2、激发性反射和M3反射。同时,也可以观察到在放上额外书的那一刻,手会有一定的下沉,但经过很短的时间手又回到原来的位置。
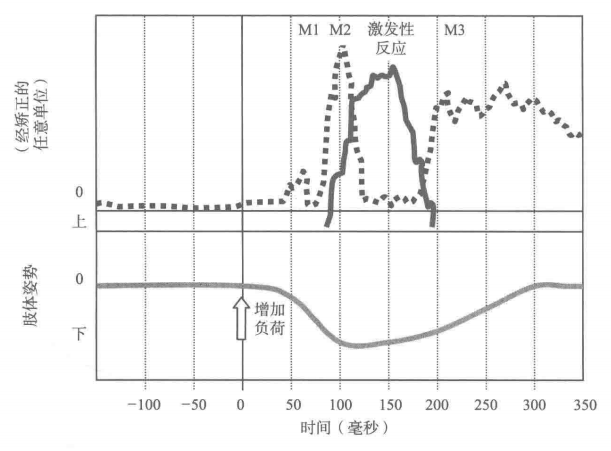
图1
二.alpha运动神经(闫剑群(主编),2015)
alpha运动神经负责肌肉的收缩,是躯体运动反射的最后公路。它既接受从脑干到大脑皮质各级高位运动中枢的下传信息,也接受来自躯干、四肢皮肤、肌肉和关节等处的外周传人的信息,许多运动信息在此会聚并发生整合,最终由它发出一定形式和频率的冲动到达所支配的骨骼肌。
要说明alpha运动神经,首先要了解骨骼肌的结构,如图2所示。骨骼肌由肌外膜包裹,其内部包含多条肌束,每条肌束由肌束膜包裹,肌束又由多条肌纤维组成,肌肉纤维称为梭外肌。alpha运动神经起源于脊髓,终止于骨骼肌。当alpha运动神经激活时,受它支配的肌肉纤维就会收缩;当alpha运动神经静息时,肌肉放松。人体的运动靠的正是这些肌肉纤维的收缩和放松进行的。一个alpha运动神经元及其所支配的肌肉纤维称为一个运动单位。当alpha运动神经元所支配的肌纤维越少时,说明相同数量的肌肉纤维拥有的alpha运动神经元越多,支配的运动越精细,运动越灵活,比如眼部运动;反之,当alpha运动神经元所支配的肌纤维越多时,则说明单个alpha运动神经元支配的运动力量越强,如小腿上面的腓肠肌。
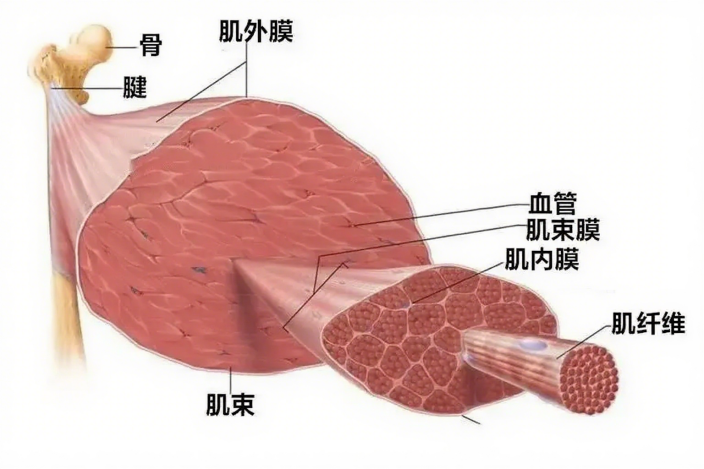
图2
三.脊髓(闫剑群(主编),2015)
脊髓对运动控制主要以反射为主,起保持身体平衡和稳定动作的作用。
1.屈肌反射和对侧伸肌反射
屈肌反射是指当皮肤受到伤害性刺激时,肢体回缩的原始保护性反射。它至少需要经过三个神经组织,首先是皮肤或关节感受器感应伤害刺激,随后感受器将刺激信号传至脊髓中枢神经,最后中枢神经激活alpha运动神经元使肌肉作出收缩反应。屈肌反射的强度与刺激强度有关,比如当低强度的刺激刺激脚的时候,仅引起踝关节屈曲,随着刺激加强,膝关节及髋关节也会发生屈曲。当强度继续加大时,这时会发生对侧伸肌反射,会引起对侧肢体的伸展,起平衡身体的作用。
2.牵张反射
牵张反射为当骨骼肌迅速拉伸而引起的收缩活动。它的基本原理是当骨骼肌和肌梭受到了牵拉,由于肌梭长度的变化会使肌梭产生反应。反应信号通过1a类运动神经传至中枢神经(脊髓或大脑),随后使对应的alpha运动神经元激活,使被拉伸的肌肉收缩(15-4:身体感觉:本体感觉)。牵张反射可分为腱反射和肌紧张两种。
腱反射是敲击肌腱时引起的牵张反射,膝跳反射(敲击膝盖下的韧带引起的)和跟腱反射(敲击跟腱引起的)都属于腱反射。因为该反射最远上传至脊髓,只经过一个中枢神经元的传递,所以反射时间很短,并且是无需意识的。肌紧张是指缓慢牵拉肌肉时产生的牵张反射。表现为被牵拉的肌肉发生微弱而持久的收缩,防止肌肉被拉长。比如人在站立时,为了抵抗重力防止关节趋于弯曲,腿上的伸肌需要持续的收缩,且背部、颈部的伸肌也需同时保持紧张,使大腿伸直,挺胸伸腰,抬头保持站姿。另外,肌梭(15-4:身体感觉:本体感觉)中关于"手持水杯倒水"的例子也属于肌紧张的一种。
3.反牵张反射
牵张反射使肌肉收缩,而反牵张反射使肌肉放松。当牵引力突然加大时,高尔基腱器受到刺激,1b类传入神经将刺激信号传至中枢神经,然后中枢神经通过反馈抑制对应的alpha运动神经元,并激活拮抗肌,使肌肉放松,从而保护肢体(15-4:身体感觉:本体感觉)。一般来说,反牵张反射只有在张力较高的时候才会被激活,而对于像"手持水杯倒水"等情况,因为肢体受到的张力有限制,这并不会引起反牵张反射。
4.间节反射
除了一些简单的反射,脊髓还能进行一些复杂的反射,如间节反射。间节反射是指由脊髓内的神经元直接产生的一种协同活动,比如双腿能协调行走。行走是一种节律运动,这种运动可以随意开始或者停止,一旦开启就无需意识参与,直到停止。中间这段无意识的节律运动就是由脊髓和脑干的中间神经元产生的,而这些神经元组成的网络被称为中枢模式产生器。
如图3,中枢模式产生器产生节间运动的原理是,在两侧肢体各自存在一个神经元,当一侧神经元被激活时,该则神经元会传递一个信号给中间抑制神经元,使中间抑制神经抑制对侧神经元,中间抑制神经元有一个自返的延时抑制信息,在一定延时后对自身进行抑制,这时对侧神经元会解除抑制(被激活),然后激活另外一个中间抑制神经元,并对该侧神经元进行抑制...。这样两则神经元交替激活和抑制,当一侧运动神经元激活时会抑制另外一侧的神经元,另外一侧的激活时又会抑制该侧的...,从而产生两侧肢体的交替运动。走路就是这样,一个腿伸,另外一个腿屈;反过来当另外一个腿伸,则该腿缩。搔爬反射也是一种间节反射,它是皮肤瘙痒时引起的一种挠痒动作。
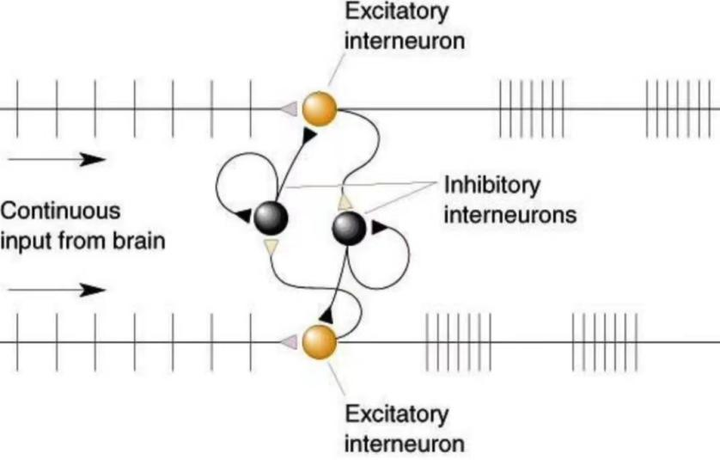
图3
一个实验证实了中枢模式发生器的存在。英国神经学家诺贝尔得主Sherrington(1947)在实验中切断猫和狗的脊髓,使大脑皮质失去了对一些alpha运动神经元和脊髓的中间神经元的控制。在对其进行适当的刺激下发现,动物身体即使没大脑信号的输入,也能进行一些类似于走路的动作。因此可以断定,这类运动模式的编码是起源于脊髓而非大脑。另外,在Brown(1911)的实验中,同时切断大脑到脊髓的下行通路和感觉到脊髓的上行通路,动物仍然可以产生完整的动作序列。这也说明了,脊髓可以在同时没有下行控制信息和没有上行感觉反馈信号的作用下产生节律运动的运动程序,如图4。
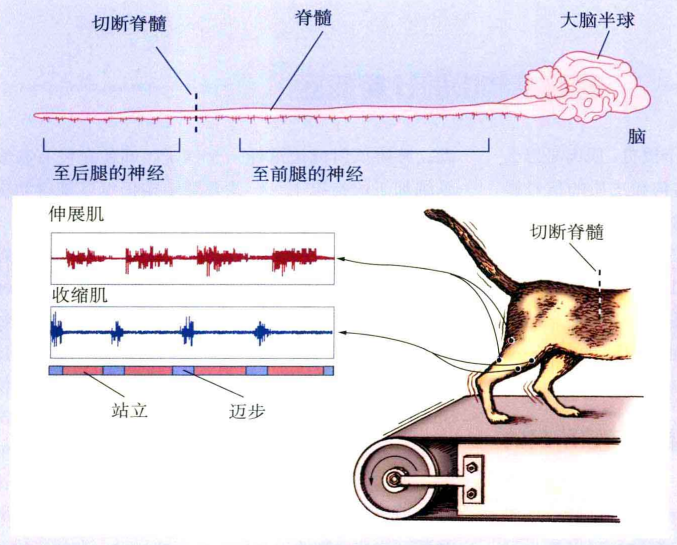
图4
四.脑干(闫剑群(主编),2015)
脑干处于高级运动神经中枢和脊髓之间,可以将感觉信号上传,高层的运动信号也经此而下行。因此它起到了一种中间桥梁的作用。
1.对肌紧张的影响
它的一个作用是通过对肌紧张的调节完成一些复杂的反射。利用电刺激脑干的不同区域,可以发现脑干中存在加强或抑制肌紧张的区域,分别称为易化区和抑制区。易化区较大,活动性较强;而抑制区较少活动性较弱。易化区略微占优。在更高位的脑区,可以发现刺激前庭核、小脑前两侧部和中间部可以增强肌紧张反射,并且这些脑区与脑干易化区有结构和功能上的联系,因此可以判断这些区域可能通过脑干易化区对肌紧张进行调节;另外,也发现大脑皮质运动区、纹状体、小脑前叶可能可以通过脑干抑制区对肌紧张进行调节。
为了研究脑干与肌紧张的关系,需要先引入去大脑僵直现象。脑干由自上而下的包括延髓、脑桥和中脑三部分,当在中脑的上下丘之间切断脑干(中断大脑皮质和纹状体与脑干抑制区的联系),会出现一种伸肌紧张增强的现象,即现四肢僵硬、头尾昂起、脊柱呈现弓反状态。当对肌肉注射麻醉或在脊髓中切断相应肌梭的1a类传入神经,这种现象便会消失。对此的解释是,因为切断的是大脑皮质和纹状体与脑干抑制区的联系,使抑制区的活动减弱,易化区明显占优。这一操作造成的结果就是伸肌的gamma运动神经元被激活,肌梭收缩,其敏感性被大大增强,从而通过牵张反射使伸肌的alpha运动神经元激活,伸肌收缩。可见去大脑僵直是由牵张反射发展而来的。当切断肌梭的1a类传入神经时,因为感受不到肌梭的长度变化,无法造成牵张反射,所以去大脑僵直现象消除。又因为这是由于gamma运动神经元引起的去大脑僵直,所以它又被称为gamma僵直。
当1a类传入神经被切断后,gamma僵直已经不可能在发生。此时,如果对小脑前叶进行切除,则可发生另外一种僵直:alpha僵直。这种僵直是由高位中枢神经或者脊髓的中间神经元作用直接作用于alpha神经元而产生的。当小脑前叶与抑制区的联系被切断,从而无法使对alpha神经元进行抑制时,被激活的alpha神经元使伸肌肌肉收缩从而产生僵直。
通过这些研究可以发现,除了脊髓的反射运动之外,还存在更高位的神经系统能直接对alpha神经元和gamma神经元进行控制。最后值得一提的是,还存在一条从脑干到中央模式产生器的通路,使大脑皮质可以对节律运动进行控制。
2.状态反射
状态反射包括迷路反射和颈紧张反射。迷路反射的感受器是前庭觉,当动物躺卧时由于前庭觉感受到重力加速度,进而使伸肌紧张达到最高的状态,相反躺卧时伸肌紧张最底。颈紧张反射是指颈部扭动时,颈部的本体感受器感受到拉伸,进而作出反射调节。如当头向一侧扭转时,身体另一侧的会产生相应的反射。
这种反射会受到高位中枢的抑制所以不容易表现出来,当去除大脑部分只保留脑干时,这种反射明显易见。
3.翻正反射
当动物被四脚朝天的从空中抛下的时候,动物能迅速的反转,变成脸朝下。在这个过程中,先是头部翻转,然后头部带动前肢翻转,最后是后肢也被带动起来,从而四肢能安全着陆。在这个过程中实际上存在一系列的反射,最初由于头部在空间中位置的不正常(空中落下),视觉和前庭觉会使头部进行翻正(脸朝下)。然后头部和身体躯干之间的位置不正常(头部不是朝着身体正前方),颈部的本体感受器会促使躯干进行翻正...。
五.运动皮质(Shumway-Cook 和Woollacott(著),刘浩等人(译),2021;Gazzaniga,lvry 和Mangun(著),周晓林和高定国等人(译),2011;闫剑群(主编),2015)
1.各运动皮质及其功能
运动皮质包括初级运动皮质和前运动皮质区,前运动皮质区又包括辅助运动区和运动前区。
(1)神经系统之间的通路
根据不同神经组织之间的连接可以区分出不同的通路,包括皮质脊髓束、皮质脑干束、前庭脊髓束和脊髓小脑束等。皮质脊髓束,顾名思义就是由大脑皮质到脊髓的通路,从初级运动区和运动前区皮质发起,经过脑干和脊髓,终止于alpha运动神经元和gamma运动神经元。皮质脊髓束可以分为侧束和前束,侧束约占皮质脊髓束的80%,控制四肢远端肌肉,与精细运动有关;前束约占20%,控制身躯和四肢近端,与姿势维持和粗略运动有关。皮质脑干束则是从皮质出发,并到达脑干的传导通路...。
(2)初级运动皮质
初级运动区可以接收来自肌肉的动觉、皮肤的皮肤感觉以及前运动皮质的输入。初级运动区神经元的输出与皮质脊髓束相连,使其既能通过gamma运动神经元控制肌梭长度,也能通过通过alpha运动神经元控制肌肉运动。这表明初级运动皮质有直接控制肌肉的能力。另外,因为初级运动区与皮肤感觉和本体感受相连,所以有学者认为初级运动区可能会在肌肉遇到意外负荷时产生额外的力量输出,从而在姿势控制中发挥作用。
早期的实验表明,初级运动皮质的细胞和脊髓中的运动神经元之间存在一对一的对应关系;然而,最新研究显示,同样的肌肉可以被多个运动皮质的神经元激活。人们还发现,大多数来自初级运动皮质的刺激会激活单一关节的多块肌肉,并作出简单运动。
(3)运动前区
运动前区的输入有小脑和顶叶等,它能将这些外源性感觉信号与动作联系起来,主要负责由外部或内部的感知觉引导的反射性运动,比如眼睛接收到苹果或者剪刀等不同的视觉刺激时,会激发不同的动作表征,如看到苹果就会联系到"拿起来吃"或者"拿起来洗",看到按钮会联系到"按按钮"...。这种感觉与动作的连结被称为联想性学习。在运动前区有病变的猴子无法将特定的刺激与对应的动作联系起来。需要说明的是,如果视觉多个目标被注意到,可能会引起很多动作表征,但最后这些运动表征最多只能有一个被选择。运动前区生成的动作表征会输出到初级运动皮质和皮质脊髓束中去。这表面,运动前区既能通过初级运动皮质间接控制运动,也能通过皮质脊髓束直接控制运动。
顶叶是处理视觉的重要脑区,从顶叶到运动前区有两条通路,分为是触及物体通路和抓取物体通路。触及物体通路通过物体在三维空间中的位置引导运动方向,抓取物体通路通过物体的视觉形状、大小等特征来控制抓握的手势(Krakauer和Ghez,2000)。另外,在运动前区中有研究表明不同的神经元控制不同的手势(Rizzolacci,Camarda,Fogassi和Gentilucci等人,1988)。
有研究表明左侧顶叶负责动作的表征和理解,生成的运动表征会输入到左右双侧的运动前区中。如果左侧顶叶受损,一方面将无法理解动作,同时也无法做与任务相关的一些动作。在一项对左侧顶叶的研究中,研究者分别让失用症患者完成两个任务(失用症是一种肌肉控制没有问题但是运动协调功能有问题的症状,发生原因可能是运动前区受损,被称为前部失用症;也可能是左侧顶叶受损,被称为后部失用症),如图5。第一个任务是运动检测,实验者让被试做一些与任务相关的动作,比如"向我展示用钥匙开门的动作"。第二个是知觉任务,让被试观看一段视频,视频中展示了三个动作,三个动作只有一个是与任务相关。然后实验者要求被试判断哪个动作属于任务的正确动作。实验结果表明,如果被试属于前部失用症,那么被试无法很好地完成运动检测任务,但他们的知觉任务的表现较好,即他们能理解知觉中动作与任务的关系,但是无法作出相应动作。如果被试属于后部失用症,两项任务均表现不佳。这是因为前部失用症只是运动前区受到损伤,健康的左侧顶叶不影响运动的视觉检测;而后部失用症属于左侧顶叶的损伤,这种损伤一方面会损害知觉能力,另一方面无法给运动前区提供运动表征,从而损害运动能力(Heilman,Rothi和Valenstein,1982)。
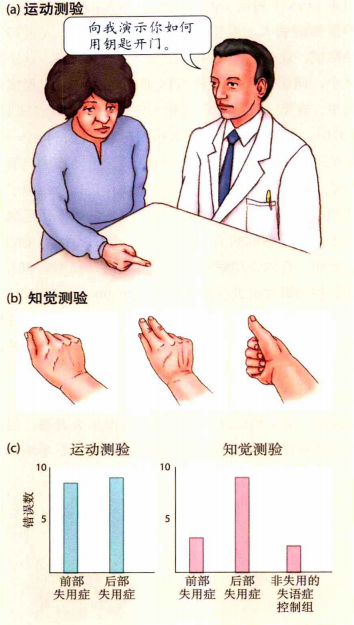
图5
(4)辅助运动区
辅助运动区接受基底神经节、额叶(目标)的输入,可以输出至初级运动皮质。处理的信息主要与运动知识、经验和自发性目标等内源性信息有关。
Roland等人(Roland,Larsen,Lassen和Skinhøj ,1980)对辅助运动区的功能进行了探索。在一个实验中,主试让被试完成一项从简单到复杂的动作任务,为了测量运动时的活动脑区,需要对被试大脑注入短效放射性注剂。结果发现当要求被试做简单重复任务时,如用拇指按按钮。在初级运动区和感觉皮质的血流量增加;当要求被试做一项复杂任务时,如四指按照顺序协调地作出一系列的动作。辅助运动区、初级运动区和感觉皮质都有血液增加;而只要求被试想象该任务而不实际执行时,那么只有辅助运动区的血液增加。可见,辅助运动区对主动的运动计划有重要作用。
有研究人员提出了运动前区和辅助运动区功能特异化的假说。Passingham等人(Passingham ,1985;Passingham,Chen和Thaler,1989)认为,运动前区和辅助运动区是根据不同的信息导向发起运动的,前者用于处理外源性感知觉引发的动作,后者负责内源自发性动作。Mushiake(1991)发现当一个运动序列是由视觉刺激引起的,运动前区更活跃,而作出一个被记住的或者自发性的动作时,辅助运动区更活跃。
人有自主行动的能力,能抑制外部刺激所引发的各种运动趋势,并对双手协调运动起重要作用。比如当环境中存在大量视觉刺激时会引发很多不同的视觉驱动运动。这时辅助运动区可以对这些运动前区发起的运动表征进行抑制或选择。对于异手综合征(辅助运动区受损)的病人,即使没有让他们行动或者让他们不要动,他们还是会用手抓握物体。进一步发现,用于抓握物体的病人的手往往是手对侧的辅助运动区受到损伤(比如,左侧辅助运动区受损,那么可以发现病患右手不受控制)。对此的解释是,运动前区由于外部刺激引发了某种运动倾向,由于辅助运动区受损无法对这种运动倾向进行抑制,所以产生了由运动前区引发的运动。
虽然辅助运动区有抑制运动前区的能力,但是人们也有可能同时依赖于内源性和外源性信息发起运动。以网球运动员为例,接球时决定打高手球还是短吊球不仅取决于接球时球到达的位置和速度(外源性),还取决于对对手能力的了解(内源性)。协调的动作是由多种来源信息指导的。
2.基本特性
(1)区域特异性
一般认为不同运动神经细胞控制着身体上的不同肌肉,而运动皮质的不同部分控制身体的不同部位,换句话说,可以把运动皮质分成很多部分,不同部分的皮质对应身体不同部位的运动控制,如左手、右脚、腰。当给予这些皮质一个电刺激时会引起相应肢体的反应。这些皮质的大小与运动控制的复杂程度有关,皮质越大,表示该皮层对身体位置的动作越复杂。手指占据着皮层的很大一部分,因此人类的手部能从事如此复杂的活动。
(2)对侧控制
大脑皮质分左右两侧,每侧大脑都是对对侧身体进行控制的。比如大脑左侧的皮层控制右侧的身体运动。在同一时刻人能做的事情是有限的,分侧控制并不意味我们能同时用两个运动表征分别控制左右两侧。如果要求双手进行不同运动任务时,两个运动可能会发生冲突从而造成干扰。比如要求一只手画圆揉胃而另外一只手上下轻拍头,这种动作是较难办到的。为了证实该理论,Franz等人(Franz,Eliassen,Ivry和Gazzaniga,1996)做了如下验证。如图6,如果要求左右手同时画一个对称的图案,那么普通人表现良好。当要求他们同时画一个不对称的图案时,被试很难画准确,如图(a)。但是对于割裂脑病人而言(胼胝体切开的病人,胼胝体是连接左右两侧大脑的通道。例如,左侧顶叶正是通过胼胝体控制右侧的运动前区),两类任务表现同样良好。这是因为胼胝体切开后左右两侧的运动任务没有干扰和冲突,如图(b)。对于健康人而言,虽然双手难以作出结构不同的运动,但是通过练习,可以把左右两侧的不同运动表征整合为一个复合表征(比如绑鞋带或打字),这样能避免了冲突,并发展出更复杂的结构不一样的运动。
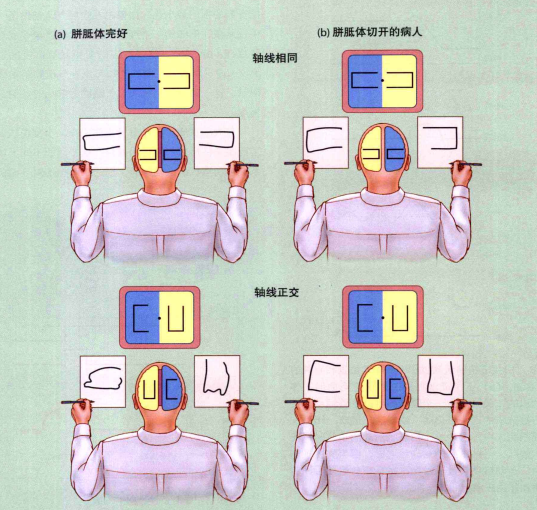
图6
大脑皮质对对侧运动进行控制,而不同的皮质之间是同侧控制。比如初级运动皮质对对侧身体进行控制。而运动前区和辅助运动区对同侧的初级运动区进行控制。
(3)层级组织和并行组织
前面提及过,对于初级运动皮质,神经元控制的往往是具体的某个关节的运动,能操作特定的肌肉。在前运动皮质区(辅助运动区和运动前区),神经元编码动作序列信息。而更高层的运动组织不负责具体的运动,左侧顶叶通过视觉刺激引导活动,它控制双侧运动前区;而额叶与运动的目的相关,它控制辅助运动区。在概念组织中论述了概念具有抽象与具体之分。运动之间其实也有这种差异,比如拿杯子,这是一种相对抽象的动作;我们可以用不同的手势拿不同的杯子,这涉及到具体的肌肉和关节,因此是相对具体的。基于此,我们看到了运动系统具有层级性,越是高层的神经组织其编码可能越抽象(最高层与目的相关),而越底层越是具体(与具体某个肌肉相关)。
Mackay(1987)提出了运动信息的层级式表征理论。该理论认为,最高级的表征是概念层,它与运动目标有关。比如图7中男士有着"想去跳舞"的意图。中间层表示运动的组织和计划,它包含实现同一个目标的多个不同的动作序列。对于该意图,实现的方式可以通过手势表达他想去,也可以口头表达"我想去"或者"我愿意去"等。最低层是运动的执行,即将这些运动计划转换为肌肉的运动。
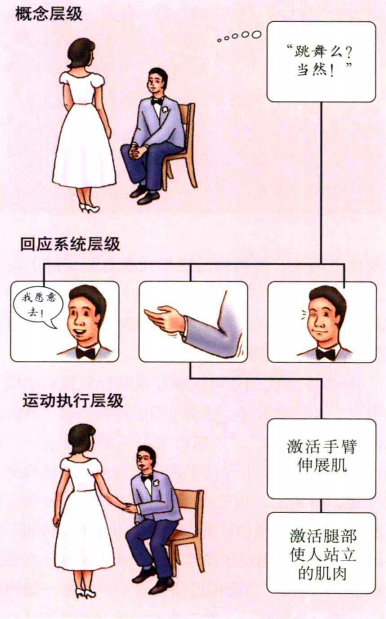
图7
我认为,不同层次表征上的注意力分布是不均匀的。一般情况下,越高层的表征能获取的注意越多,反之则越少。很多时候对于熟悉的动作,除非我们刻意的去注意具体的肌肉或关节运动(底层),否则注意可能更多的停留在刺激或者目的层面(高层)。比如在写字时,我们更多的是注意我们要写的那个字,而非注意具体的笔画。我们很少会去想(注意)"王"字的几个笔画,"横横竖横"四笔每一步的操作,而只需要想"写王字"这个意图,然后一切就很自然会进行下去。在非新手打羽毛球、乒乓球等球类运动时,注意通常也只在目的层面和刺激层面。一般玩家很难说出他们是如何利用肌肉把球接住的,因为更多的注意是停留在球的运动轨迹或者需要接球的目的。又如用乐器弹奏一首曲子,在熟练的情况下对肌肉控制的注意往往很少。当然,我们也可以特意去控制特定的肌肉运动(通过注意去制造这样的目的)。这说明,我们在发出动作时更多关注的是刺激或者目的,对于熟练动作,高层表征所分配得的注意会比底层表征的多。
除了分层组织,运动控制还通过并行组织来处理运动信号,分层组织和并行组织共存。从上面内容中我们已经看到了并行控制的例子。运动前区、初级运动皮质和脊髓都有直接控制肌肉的通路,使一块肌肉即能自主控制(通过初级运动皮质或运动前区直接控制),又能被动控制(通过脊髓的反射);另外一个例子是,不同的视觉刺激可以同时在运动前区引起不同的运动趋势,同时辅助运动区又能产生自主性的行为。
(4)冲突处理
动作可以由不同层级的神经系统产生,如底层级的脊髓反射动作和高层级的运动皮质控制动作。高层级的神经组织一般能对低层级的动作形成抑制,如高级的意识可以抑制一些低级的反射动作(如状态反射)。
运动前区主要负责刺激驱动的运动,辅助运动区负责自发性驱动的动作,因此同一种情景下,刺激驱动的运动和自发性驱动的运动可能会发生竞争。在Wise等人(Wise,Di Pellegrino和Boussaoud,1996)的一个实验中,在正式实验前会训练猴子如何进行实验。实验先给猴子呈现一个刺激,经过一段延迟时间后发出"go"信号,猴子在收到"go"信号后根据呈现刺激摆动摇杆。这个实验有两个实验条件(如图8),一个是一致性条件,当八盏灯的其中一盏灯亮了时,猴子操控遥控杆从中心位置到到灯亮的位置;另一个是不一致条件,猴子操纵遥控杆从中心到灯亮的反方向位置。实验记录了猴子的脑电信号。在一致性条件下,脑电信号在go之前在运动方向上被一定程度上的激活,go之后在运动方向上增大。这是因为刺激方向与运动方向一致,运动前区出现了由视觉引导的运动计划,如图9(a)所示;在不一致条件任务中,脑电所记录的运动表征在go之前是指向的刺激方向,然后在go之后才转向相反的方向,如图9(b)所示。这是因为产生了两个运动计划,它们之间相互竞争,一个是由运动前区产生的视觉引导运动,另一个是由自发性目的引导的运动。一开始视觉引导运动占主导,然后是目标引导运动占主导。一个动作可以被分成一个动作序列,而冲突可能仅仅发生在序列的某些部分上。在这个实验中,动作大致可分为手握遥控杆,然后才是往某个方向移动,对于这个实验的不一致条件,不论是刺激驱动动作还是自发性驱动动作,它们都包含握遥控杆,而冲突的部分仅仅是往某个方向移动,因此在握遥控杆时不会有冲突感。
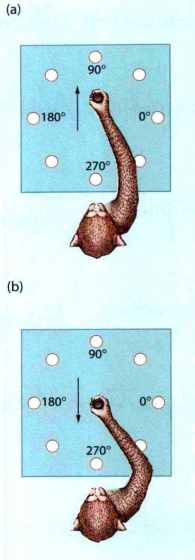
图8
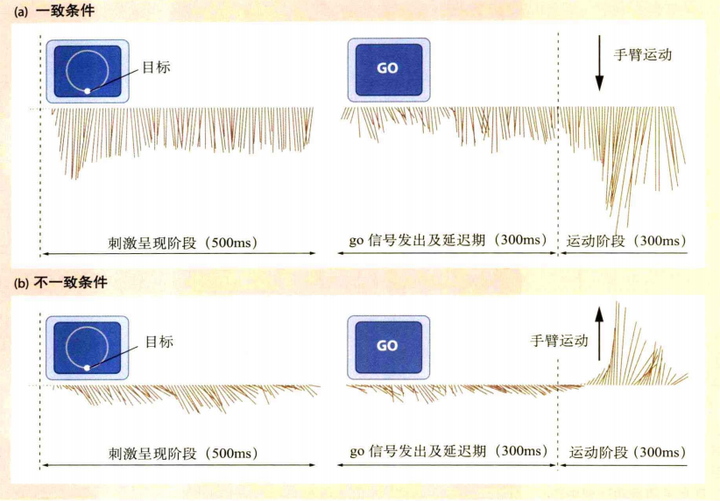
图9
辅助运动也会同时通过胼胝体对对侧辅助运动区及对侧初级运动皮质输出抑制信号。在一类辅助运动区的脑损伤研究中,要求患者使用脑损伤部位对侧的手进行某一运动时,患者同侧的手也会作出一个类似的动作(即使没要求同侧的手运动)(Wiesendanger,Rouiller,Kazennikov和Perrig,1996)。如要求用一个手拉抽屉并用另外一只手拿东西,患者会作出两只手同时拉柜子的动作。这是因为,一侧的辅助运动区在对对侧运动进行控制的同时,会抑制同侧的同类运动。如果某侧辅助运动区受到损伤,由于无法抑制同侧运动,两只手会倾向于作出相同的运动。辅助运动区对对侧运动的抑制是重要的,它使我们能作出一些双手协调的动作,如绑鞋带或钢琴演奏等。
Rosenbaum等人(Rosenbaum,Slotta,Vaughan和Plamondon,1991)就提出过一个皮质选择假设。例如,完成一个目标可以使用多种不同的方式,比如拿一杯咖啡,即可以用左手拿,也能使用右手拿,或者侧一下身体使手与杯子的距离更近。另外,如果杯子旁边有一支笔的时候,在视觉刺激驱动下可能也会产生拿笔的额外表征(即可能产生与目标无关的运动计划)。多种运动表征之间形成竞争关系,但最后只会有一个被选择。影响选择的因素有很多,比如哪个动作是利手的、对动作的习惯程度和熟悉程度、根据经验预测得出哪个动作更可能完成任务等等。最后,初级运动皮质接收前运动区和辅助运动区的输出,它可以看作一个计分器,在所有运动计划中得分最高的将被选择,最后对肌肉输入控制信号。
上面提及到运动竞争涉及利手、习惯和熟悉程度等等。而我认为还有一个重要因素:注意。试想想,当刺激驱动动作和自发性驱动动作发生竞争时,自发性驱动动作多数情况下总是能胜出。显然,自发性驱动动作的动作计划是"占据"大脑的,拥有更多注意。我们能根据我们的目的作出任何动作(只要我们能做到),比如我们金鸡独立或者空翻,没有任何刺激驱动我们这样做(相反我们的身边每时每刻总是存在各种各样的刺激驱动我们做其他动作),我们能这样做的原因只是因为我们想或者我们需要,而当我们"想"的时候被想的内容就占据着我们的注意。因此,注意在运动竞争中有重要作用。
我们身边总是有大量信息,而这些刺激都包含各自的刺激-动作联想。比如看到门把手就联系到开门和关门、看到按键就联系到按按钮等等。但是在同一时刻,能被视觉注意到的信息是很有限的(9-15:视觉注意)。因此,在一个视觉画面中,只有被注意到的数量有限的刺激能激活对应的动作。有一些刺激很容易会引起注意(刺激驱动注意),如警车上的警灯。某些时候在马路上走且注意不在路面的时候(如看手机),一些行驶的车辆更容易吸引注意,从而使行人止住脚步。有时,刺激驱动的运动甚至能压制过自发性驱动。比如玩蹦极,很多人因为高度紧张而不敢往前再走一步。越是有往下跳的趋势就越害怕,从而越被吸引注意。总得来说,刺激驱动的动作也能"战胜"自发性动作,关键要看该刺激在多大程度上驱动注意。
(5)编码内容
一个动作所包含的编码类型可能是多种多样。比如弹一首钢琴,需要匹配每个手指与按键(手指的位置编码),每个按键的力量(力量编码),将其组成序列(动作序列编码),还有弹奏的节奏(序列的时间编码)。多种编码方式为控制过程提供额外的灵活性。
(a)力量与速度
Evarts(1968)在猴子进行腕部运动时记录下了其皮质脊髓束的活动情况,发现皮质脊髓束的放电频率与运动中力量的大小以及力的变化率有关。这表明,初级运动区与肌肉的力量和运动的速度有关。
(b)位置编码
除了在初级运动区和皮质脊髓束中发现了编码力量和速度相关的神经元。事实上,大脑皮质对运动表征的编码是广泛存在的(包括运动皮质、神经基底节、小脑和上丘等)。那么,这些动作编码都包含哪些内容呢?当伸手拿一个东西的时候,其动作表征可能是手到物体的运动轨迹,也可能是运动的方向,也可能是物体所在的位置。在Bizzi等人(Bizzi,Accornero,Chapple和Hogan,1984)的一项研究中发现,相比其他表征,位置表征可能更加占优。在研究者的实验中,场景是一个漆黑的房间并有几盏熄灭的灯,当实验开始时其中一盏会亮起然后又熄灭,在灯熄灭后需要被试猴子把手伸向原来亮灯的位置。实验的关键点是,首先,在猴子发出动作后人为的对猴子胳膊施加一定的反作用力,阻碍手臂继续运动;其次,猴子的感觉传入神经是被阻滞的,而且房间是黑的,因而猴子并不知道它们的手被阻碍了。研究者认为,如果动物知道肌肉的爆发会将胳膊带走一段距离,施予反作用力会使动作终止在离目标位置还有一般距离的地方。但是如果动物产生了一个指定于目标位置的动作指令,当外力撤销时它仍会完成这一目标。实验结果支持了编码目标位置的假设。
(c)方向、距离和轨迹
位置编码是运动控制的基本能力,但并不代表运动细胞只能编码位置。事实上同样的运动可能会有多种表征,比如手臂伸直这个动作,该动作表征即可以是手的终点位置,也可以是手的位移方向和距离。在舞蹈等一些体育运动中,运动轨迹比起终点更加重要。
(d)运动序列
Kakei等人(Kakei,Hoffman和Strick,2001)对运动前区的测试中发现这里几乎所有的细胞都编码了运动方向。而另一项研究则表明,对于运动前区和辅助运动区,虽然这两个区域的细胞与运动方向有关,但也只是在特定的运动序列中才会有较强反应。而其在他序列中,同样的动作可能只会有微弱反应。这说明,这两个运动皮质与动作序列有关。
(6)表征形式
Georgopoulos等人(Georgopoulos,Kalaska,Caminiti和Massey,1982)发现,表征方向的运动神经元并不是对某个精确的方向进行表征,它表征的是一个模糊的方向范围,当肢体向某一片连续的方向运动时,该神经元都会被激活。通过测量,每个细胞的反应频率与运动方向的关系呈现如高斯分布的钟形曲线。即一个细胞对一个方向最为敏感,放电率最高,然后沿方向周边逐渐递减,如图10(a)。
一个精确的运动可能由一组运动神经元负责。表征动作的细胞并不是如祖母细胞理论一样,即一个细胞表征一个精确方向。而研究支持的是细胞以分布式编码的方式表征动作,并且该动作表征等于各细胞向量之和(每个细胞可以表示为一个向量)。以表征方向的运动细胞为例。因此,可以把该细胞的表示为一个向量,向量方向为实际的运动方向,向量长度为神经元在该方向的放电频率。当实际运动方向和神经元的表征方向(最敏感方向)越接近时,放电频率越高,向量长度越长。如果一个神经元表征的方向是模糊的,那么可以使用一组神经元表示。当要表征某个精确的运动方向时,可以把一组细胞的向量之和求得这个运动的运动方向。例如,在图10(b)中使用两个方向神经元(向量1+向量2)表征一个运动方向;而在图(c)中,显示了多个神经元如何表征不同的运动方向。Georgopoulos将这种表征称为场向量。这种运动表征更加鲁棒。
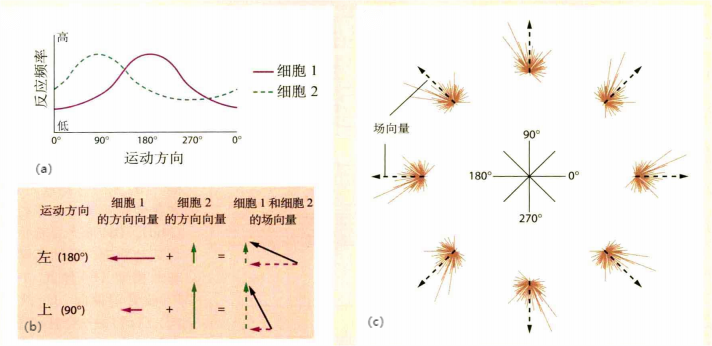
图10
(7)运动皮质的学习
Jenkins等人(Jenkins,Brooks,Nixon和Frackowiak等人,1994)进行了运动学习与皮质代谢之间关系的研究。实验要求被试学习一个包含8对声音-动作的序列。每种声音对应1个按钮,一共四种按钮,当被试听到一个声音时需要按下一个按钮。一开始被试只能靠试,每按对了就会有提示,按错了没有提示。被试会对该序列进行试错并练习。当学会这个序列之后,实验会要求被试以相同的程序学习新序列。通过对被试脑电扫描的情况可知,在进行旧序列时,辅助运动区的激活情况更强;而在学习新序列时,运动前区的激活更强。可以看出,在学习的初期,主要靠感觉信息引导学习;而在学习后期,可以靠目的引导,其依靠感觉信息的程度会比学习初期的少。这种原则在生活中随处可见,如在学习键盘打字的初期,需要用视觉对键盘进行逐个对照,当能熟练打字的时候就不需要过多的依靠视觉。另外,还有研究表明,当运动序列被熟悉后,对该运动序列的控制可以转移至初级运动皮质(Krakauer和Ghez,2000)。
大多数运动都有两类反馈,一类是外源信息(如视觉、听觉),另外一类是内源信息(如平衡觉和躯体感觉)。这两类信号对于运动的作用是不同的。在运动学习的初期,主要依靠外源信号的反馈来调整动作。当熟练到了一定程度时,可以通过内源信息来调整动作。比如对于网球新手和网球运动员来说,新手通过发球后观察球的弧线或落地点来调整动作;而运动员则可以在直接发球时通过对动觉来调整这些,还如经常打篮球的人在投球出手的瞬间就知道能不能进球,这些更多的是依靠对内源信息来预测的。通过学习,这些内源信息与运动建立了联系,从而作为内源性预测的基础。
运动表征具有不同的层级,那么学习是否可以发生在不同的层面上呢?答案是肯定的。比如一般人使用右手学习写字。但是当我们学会用右手写字时,我们还可以用左手、脚、牙齿来写,这表明有更抽象的表征来控制这些肢体运动。有研究认为,当学习了一个新动作时,学习可能发生在更为抽象的层面,建立刺激-动作联结或目的-动作联结,将动作序列组织起来等等。当熟练动作时,学习主要发生在肌肉上,比如掌握力度和节奏等。
(8)大脑皮质的反射运动
大脑皮质还能参与一些反射运动,包括跳跃反射、放置反射和抑制肌紧张。跳跃反射是指身体受到推力时,为了维持身体平衡产生跳跃的动作。把动物眼睛蒙住并且悬吊在空中时,当让动物任何一部分接触一个支持面,动物马上会用它的两只前爪放在该支持面上,这种反射叫放置反射。
六.小脑(Gazzaniga,lvry 和Mangun(著),周晓林和高定国等人(译),2011;闫剑群(主编),2015)
小脑可以分为前庭小脑、脊髓小脑和皮质小脑。不同的小脑部分负责不同的功能。前庭小脑负责身体平衡及其他一些反射运动,是进化中最早出现的,在小脑的最内部。脊髓小脑负责运动过程中的协调,它附加在前庭小脑外。皮质小脑负责存储运动程序和运动学习等高级功能,它是最晚出现的,在小脑的最外层。如图11。
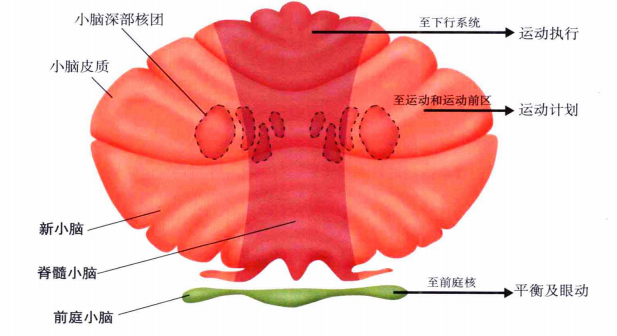
图11
1.前庭小脑
前庭小脑主要由绒球小结叶构成,前庭小脑主要通过三种反射控制身体运动,分别是前庭眼动反射、前庭脊髓反射和前庭颈部反射。其神经通路是,首先脑干中的前庭核接收来自前庭器官的前庭觉信息。而前庭小脑与前庭核有双向的联系,即接受前庭核的前庭觉信息,又对前庭核输送运动控制信息。最后,前庭核可通过内侧纵束控制前庭眼动反射;同时又可通过前庭脊髓束(从前庭核到脊髓的通路)到达脊髓控制躯干和四肢近端肌肉,以调节的身体平衡,如图12。
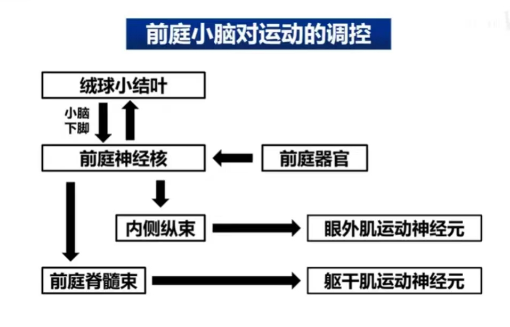
图12
前庭眼动反射,即当头部转动时,前庭器官的半规管受到刺激,而前庭小脑会将平衡觉信息转换为双眼的运动,眼部运动方向是头部运动的反方向,使眼睛能对物体进行注视。比如当头部水平左转时,双眼会有一个沿水平向右边的转动。这个转动是自动化的。正如一般相机对运动物体拍摄时都会有一定程度的模糊一样,我们运动时看物体也会模糊。但因为在眼动反射的存在,我们的眼睛有一个与头部运动方向相反的转动,所以我们始终能对物体进行注视,从而看得更清。眼震颤是一种前庭眼动节律运动,分为慢相动和快相动,两种相动交替进行。当头部转动时,眼睛会沿着运动方向的反方向慢速运动,这是慢相动。当眼睛移动到最边的一端而无法移动时,眼睛会快速回到眼中央,这是快相动。前庭脊髓反射(又称为紧张性迷路反射或躯干四肢紧张性迷路反射),它以前庭觉为感受器,身体为效应器,目的在于保持身体的平衡。比如,当汽车加速前进时,身体由于惯性有往后倾的运动趋势。但因为椭圆囊受到刺激触发了反射,躯干的屈肌会加强,从而保持身体平衡。又如,升降电梯刚刚上升时,由于加速度会刺激前庭觉,四肢会出现反射性的屈曲。而电梯刚刚下降时,下肢会伸直。前庭颈部反射,指的是身体在运动过程中为了维持头部稳定的反射。
2.脊髓小脑
脊髓小脑包括蚯部和半球中央部。如图13,蚯部下行至脊髓前角内侧(控制躯体近端的运动),也上行至负责躯体近端区域的运动皮质,因此蚯部主要与姿势维持和粗大运动有关;半球中央部下行至脊髓前角外侧(控制躯体远端的运动),也上行至负责躯体远端的运动皮质,因此半球中央部主要与精细运动有关。另外,脊髓小脑也接收初级运动皮质从皮质脊髓束侧支传入的运动信息,动觉、视觉和听觉等反馈信号也会传至脊髓小脑。其中,动觉通过脊髓小脑束直接传入到脊髓小脑。
脊髓小脑的主要功能是调节正在进行的运动,以减少运动偏差。首先,初级运动皮质在进行运动控制时,其运动计划会有一个副本传入脊髓小脑(通过皮质脊髓束侧支)。动觉、视觉和听觉等对这个运动计划的运动执行进行感知,同时将感知信号上传至脊髓小脑。运动计划的副本与这个感知信号会在小脑中计算它们之间的误差。这个误差一方面会上传至运动皮质,以修改运动计划;另一方面,这个误差也能直接到达脊髓修改运动误差,使运动能按目标进行。如果脊髓小脑损伤,失去了纠正偏差的能力,其动作会变得非常粗糙和不准。比如拿杯子,在伸手的过程,要么伸过杯子,要么够不着。在每次够杯子的时候,会在超过和返回杯子的过程中来回进行,总之无法准确定位目标。同时,越是接近目标,会越容易出现震颤的现象,称为意向性震颤。另一种脊髓小脑损伤的症状是在行走的时候,要么腿迈得老开,要么迈不开。
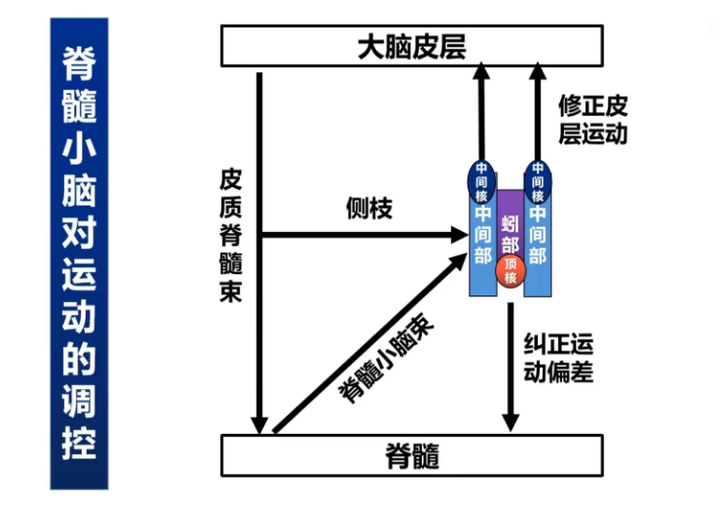
图13
最后,脊髓小脑能对脑干的易化区和抑制区进行控制,进而控制肌紧张。
3.皮质小脑
皮质小脑从脑桥(脑干的一部分)中接收来自感觉皮质、运动皮质和联络区大脑皮质的信号,并最终返回到运动皮质中。这条通路的作用是学习运动和存储运动程序(随意运动)。皮质小脑正如一个图书馆,里面存储着各种各样的运动程序。当大脑需要发起某种运动时,如弹钢琴、跳舞、打字等精细动作,可以先从皮质小脑将该运动的运动程序提取出来,并传至运动皮质,再由运动皮质控制肌肉进行该运动。这些运动程序并不是先天就存储在皮质小脑的,而是通过后天对动作的反复操练形成的。脊髓小脑可以用于比较运动计划与运动执行之间的差异,并在运动过程中逐步调整动作,使整个运动序列协调起来。在这个过程中,小脑与大脑皮质存在大量的交互,使运动程序被保存于皮质小脑中。总结来说,在学习某一动作时,脊髓小脑负责协调运动过程中的各个子动作。而动作学会以后,运动程序能从皮质小脑中提取并直接用于运动,不需要重新协调。对于皮质小脑受损的人,因为存储不了新的动作程序,所以每次练习动作时,都像第一次学习的一样。
关于小脑如何学习或者学习什么有两种假说。一是时间安排和节律控制假说,二是运动预期假说:
(a)时间安排与节律控制假说
小脑被认为具有表征时间信息的能力(这种表征还可以被用于其他系统)。而时间和节奏对运动来说是很重要的,比如棒球运动中,只有在正确的时间挥棒才能击中并将球打到指定位置;同样的,节奏对于乐器演奏也是重要的,如果节奏不对,即便每个音都对,也会影响整首曲子。Perrett等人(Perrett,Ruiz和Mauk,1993)研究了小脑与时间安排之间的关系。如图14所示,在实验中使用气流喷射器对准兔子的眼睛喷射。由于气流对眼睛产生了刺激,因此兔子会闭眼。在该实验中,实验者添加了一个程序,在喷气前会给出一个声音刺激。如果兔子通过练习习得声音与气流之间的时间关系,那么它就能准确的从声音中预测到气流的喷出,从而闭上眼睛。实验结果表明,正常的兔子能很好地预测声音与喷射器喷射气体之间的时间关系,而小脑损伤的兔子则不能。
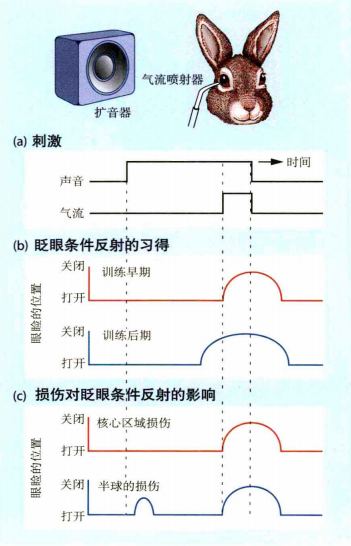
图14
在快速运动中主动肌和拮抗肌的协调运作是很重要的,主动肌负责驱动肌肉运动,拮抗肌负责制动。另外,主动肌和拮抗肌的时间安排还需非常精准才能做出正确的动作。在一个实验中,分别对正常被试和小脑损伤被试进行快速挥手测试。正常被试能正确预测拮抗肌的制动时间,主动肌和拮抗肌的启动终止时间控制精确;而小脑损伤被试的拮抗肌出现了延时启动,主动肌和拮抗肌的时间安排出现了混乱。
当小脑损伤时还可能会发生运动分解缺陷,即原本同时发生的动作会变为顺序发生。比如伸手拿东西的时候,正常情况下在手接近物体的同时也会作出张手的抓握动作,如图15(a);而在动作分解缺陷的情况下,抓握的动作并不是在伸手的过程中发生的。而是触碰到物体时再抓握,是顺序发生。从而影响了协调性,如图15(b)。
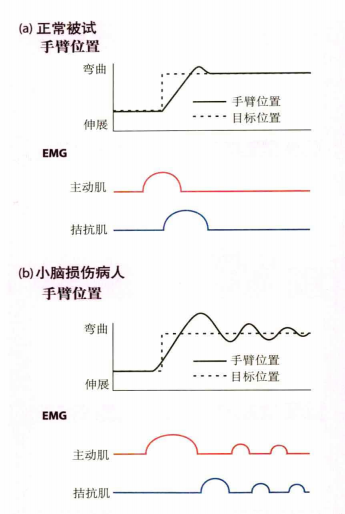
图15
(b)运动预期假说
运动预测,可以通过运动与运动产生结果的联结从而产生预测。这里可以分为自我运动和其他运动。自我运动是对自身运动时间空间轨迹的预测,比如熟练的网球手能够将球击到某个位置,或者在击球的一瞬间能够预料到球是否打偏。又如下楼梯时脚踏空了会感觉非常惊讶(因为和踏上阶梯的预期不符)。其他运动是对其他物体运动的时间空间预测,比如只有预判球飞来的时间和位置,球员才能更好的接球。在过马路的时候常常会有这种体验,当车距离较远或者车速较慢时能放心的过马路,而车较近或者车速较快时注意力会高度集中。这里启动了对自我和车运动关系的预判。
在一些运动学习的脑成像研究中发现,在学习一个新技能的时候,小脑一些区域的活动会在练习新技能时增强,而当动作逐渐熟练后这些区域的小脑活动逐渐减弱。据研究猜测,这些脑部活动可能是由于对运动的预测导致的(Imamizu,Miyauchi,Tamada和Sasaki等人,2000)。
七.基底神经节(Gazzaniga,lvry 和Mangun(著),周晓林和高定国等人(译),2011;闫剑群(主编),2015)
1.直接通路、间接通路和超直接通路
基底神经节由尾状核、壳核、黑质、苍白球、丘脑底核五个核团构成,苍白球又可分为内侧和外侧两部分,而黑质包括黑质网状部复合体和黑质致密部。
(1)直接通路和间接通路
尾状核和壳核被称为纹状体,它是基底神经节的输入区。大脑皮质广泛区域(感觉区、运动区、联合区等)的信息都是由纹状体进入。苍白球内侧部分和黑质网状部复合体是基底神经节的输出区,基底神经节所有的信息都是由这里传出。最后,输出信息再通过丘脑回到大脑皮质运动区。基底神经节包括输入连接,内部连接和输出连接三部分。广泛皮质区到输入区的通路是输入连接,其输入是兴奋性的(即信号可使输入区激活,从而使其活动增强)。从输出区到丘脑是输出连接,其输出是抑制性的(即信号可使丘脑抑制,从而使其活动减弱)。丘脑到各大脑皮质的信号是兴奋性的。内部连接则是五个核团之间的连接。如图16所示。
从输入连接到内部连接再到输出连接包括直接和间接两条通路:
(a)直接通路:直接通路是"皮层-纹状体-输出区-丘脑-皮层"的通路。纹状体到输出区是抑制性的,从输出区到丘脑是抑制性的。当广泛皮质区给纹状体输入兴奋性信息时,纹状体活动性增强,然后对输出区发送的抑制性信息就会增多,输出区受到抑制时就会减少对丘脑发送抑制信号,因此丘脑活动性增强,从而易化运动皮质。在这里,通过两次抑制最终得到兴奋作用(负负得正)的现象叫去抑制。相反,当纹状体没有任何输入,纹状体的活动性就会减弱,因此给输出区发送的抑制性信息减少,然后输出区的活动性增强,然后输出区给丘脑发送的抑制性信号就会增多,最后丘脑就会减少给各皮质发送的兴奋性信号,从而抑制运动。
(b)间接通路:间接通路是"皮层-纹状体-苍白球外侧-丘脑底核-输出区-丘脑-皮层"的通路。纹状体到苍白球外侧是抑制性的,苍白球外侧到丘脑底核是抑制性的,丘脑底核到输出区是兴奋性的。根据之前对直接通路的分析,便很容易得出间接通路的运行模式。通过分析,直接通路和间接通路互为拮抗。当大脑皮质对纹状体发送兴奋性信息时,在经过直接通路后可促使丘脑对运动皮质发送兴奋性信号,而经过间接通路后则是抑制运动皮质。它们之间的冲突并非同时发生。可以看出,间接通路比直接通路多两个节点,因此间接通路的通行时间比直接通路的长,可以在运动过后起到制动的作用。还有一种说法是,直接通路只激活特定的神经元,而大部分神经元会被抑制,因此特定的动作从众多可能动作中被选择。
(2)黑质致密部-纹状体
这里还需要提到从黑质致密部到纹状体的通路。黑质致密部有两种类型,一种包含D1受体,它将信号发送至纹状体内侧并产生兴奋性作用,控制直接通路;另外一种包含D2受体,它将信号发送至纹状体外侧产生抑制性作用,控制间接通路。虽然D1受体与D2受体传递的信号不同(一个兴奋性,一个抑制性),但是它们对大脑皮质的效果是相同的。因为通过分析可知,D1到丘脑的直接通路和D2到丘脑的间接通路到最后都是易化大脑皮质和促进运动的。D1受体与D2受体可能分别与不同的任务相关。Nonomura等人(Nonomura,Nishizawa,Sakai和Kawaguchi等人,2018)用光激活D1受体时发现了大鼠倾向于重复原来的动作,激活D2受体时则倾向切换动作。
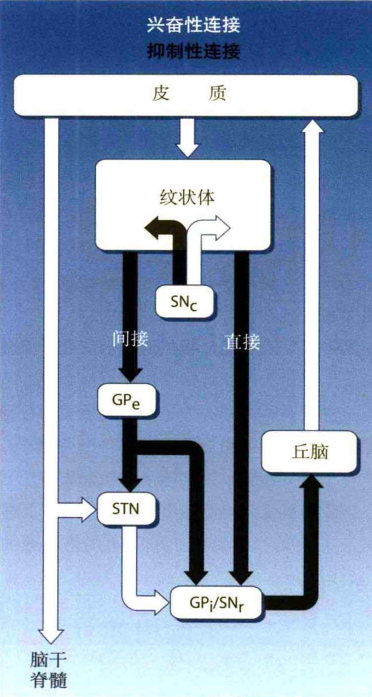
图16
(3)超直接通路
还有一种超直接通路被提出,即"皮层-丘脑底核-输出区-丘脑-皮层"的通路。兴奋性信息从大脑皮质区出发,然后直接进入丘脑底核,使丘脑底核活性增强。在此通路中,丘脑底核与输出区有着广泛的纤维投射。从而使输出区被广泛增强。最后,再从丘脑到大脑皮质,使大面积的大脑皮质被抑制(Nambu,Tokuno,Inase和Takada,1997;Nambu,Tokuno和Takada,2002)。
2.基底神经节的功能
(1)运动的起止控制
对于基底神经节作用的一种说法是控制随意运动的动作起止。近年来,研究者对超直接通路、直接通路和间接通路进行研究。三条通路到达输出区的传递时间分别为7.8 ms,20.9 ms和29.9 ms。按照时间的先后顺序,这三条通路使运动先后经历"大面积抑制-增强-抑制"三个阶段。在大面积抑制阶段(对应超直接通路),其他刺激引起的或其他不必要的动作会被抑制,它的作用相当于运动的预备阶段,屏蔽干扰;在增强阶段(对应直接通路),对应的运动计划被执行,进行动作;在抑制阶段(对应间接通路),被激活的运动计划在所在的脑区被重新抑制,运动终止(Nambu,Tokuno,Inase和Takada,1997)。
(2)运动的选择
Albin等人(1989)发现皮层-基底神经节环路存在着大量的并行分支、反馈环路和互惠连接的附加连接,这提示基底神经节可能属于更复杂的模型。一些研究者认为,直接通路和间接通路并非简单的起止关系。而是直接通路主要负责目标动作的发起,而间接通路则负责对其他动作的抑制,以防止这些动作对目标动作造成干扰(图17)(Cui,Jun,Jin和Pham等人,2013;Surmeier,2013;Sheng,Lu,Shen和Poo,2019)。
这种多通路的方式能很好的解决运动竞争的问题。直接通路中,特定的运动计划只会使特定的神经元激活;而其他通路则继续保持对各种可能动作的抑制。从而能从各种备选动作中提取一个动作,做到赢家通吃。
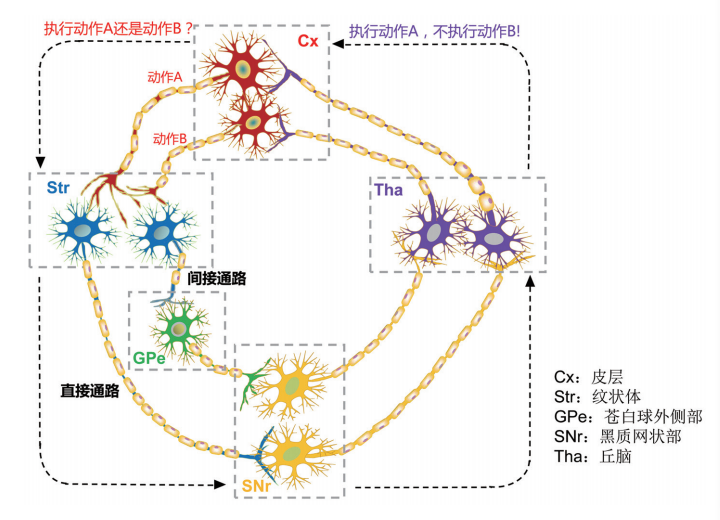
图17
(3)转换假说
一些研究发现,基底神经节对心理定势的转换有关键作用。Hayes等人(Hayes,Davidson,Keele和Rafal,1998)开发了运动转换和认知转换两种任务来研究该问题。在运动转换任务中(图18(a)),实验者训练被试完成两个动作序列,如用手指按"I-R-M"和"I-M-R"。之后被试需要在两种条件下进行测试。非转换条件下,被试需要完成两组相同的序列,如"I-R-M-I-R-M"。在转换条件下,被试需要完成两组不同的序列"I-R-M-I-M-R"。实验结果表明,正常被试在两种条件下的成绩都很好。而相比非转换条件,帕金森氏症的病人在转换条件下,动作会显著变慢。在知觉转换任务中(图18(b)),需要被试判断颜色或者形状,一次试验包含两次辨别。该实验同样包含两种实验条件。在非转换条件中,两个辨别任务相同(如颜色判断-颜色判断)。在转换条件中,两个辨别任务不同(如颜色判断-形状判断)。结果显示帕金森氏病人在转换条件下的表现会更慢。因此,不论是动作转换还是知觉转换,都需要基底神经节的参与。除此之外,从抽动症和强迫症(与基底神经节有关的疾病)病患的身上,能看到一些重复的刻板运动。其中,抽动症是指简单的抽搐、耸肩等动作,强迫症比如"不停的洗手"等动作。
上述这些现象都在一定程度上说明了基底神经节的转换假说。神经基底节监督各皮质的活动,通过撤销特定神经元的抑制,为不同心理定势的转换提供可能。
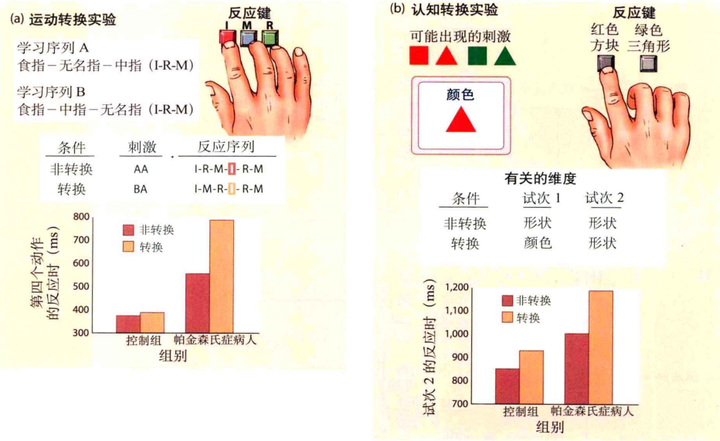
图18
(4)运动学习
基底神经节还与运动学习有关。黑质致密部(包含D1或D2受体)对纹状体输送的信号是一种多巴胺。多巴胺在大脑学习中扮演重要角色,通过提供奖励强化成功行为与对应环境的联系。当需要适应不熟悉的环境或者打破熟悉环境的习惯时,在多巴胺的释放和动作的成功执行后,多巴胺会改变基底神经节的通路。往后如果动物处于相同(相似)的环境下,动物更倾向于执行之前成功过的行为。最后通过学习,动物从不同的环境中学习到各种动作。而对于每种环境与每种动作的组合,多巴胺有着不同的强度。
(5)运动编码
最后,基底神经节还参与了运动速度、运动方向、运动强度等运动编码的调控(黄浩洁,2019)。
3.亨延顿氏舞蹈症和帕金森氏症
当基底神经节病变时,有可能会产生两种运动性疾病,亨延顿氏舞蹈症和帕金森氏症。亨延顿氏舞蹈症的主要症状包括不停的运动,失去平衡,笨拙、姿势扭曲等。正常运动逐渐被非正常运动支配。尸检发现,这类病人的基底神经节萎缩明显,且纹状体90%的细胞处于死亡状态。在间接通路中,从纹状体到苍白球外侧的抑制力变弱。使最后的输出连接对丘脑的抑制力减弱,造成丘脑对运动皮质的兴奋性信息增多(图19(a))。帕金森氏症的主要症状包括全身肌肉紧张、肌肉强直、随意运动减少、动作缓慢、运动功能减退等。帕金森氏症的病人在改变姿势上比较困难,当发起新动作时这种问题更加明显。帕金森氏症主要与黑质致密部的病变有关。失去了黑质致密部的两种信号(D1和D2)的影响,致使直接通路的兴奋性信号减弱,间接通路的抑制性信号增强,最后使输出区对丘脑的抑制增强。与亨延顿氏舞蹈症相反,帕金森氏症的大片运动皮质受到抑制(图(b))。
这两类病变实际上都属于自主运动的丧失,亨延顿氏舞蹈症很难对运动进行抑制(包括各种感觉驱动的运动),它是各种运动表征混合和冲突的结果;帕金森氏症很难随意运动,正常人在没有运动计划或不执行运动时,肌紧张占主导,负责起姿势稳定和平衡的责任。当在进行随意运动时,运动皮质的随意运动表征占主导。由于帕金森氏症的随意运动信号被抑制,导致肌紧张过强。
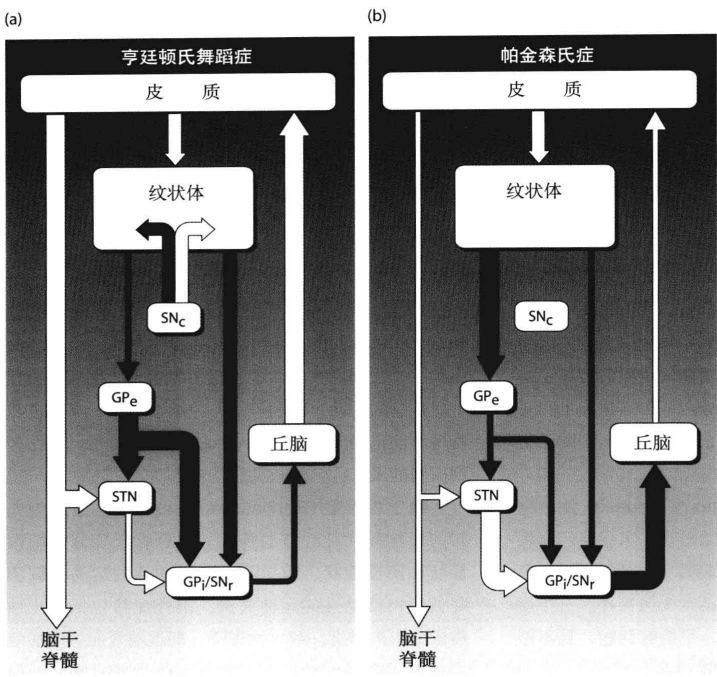
图19
4.神经基底节与小脑的关系
小脑和基底神经节同属皮质下区域(大脑皮质外的脑区),它们与运动皮质构成了最高水平的运动控制系统。与初级运动皮质不同,小脑和基底神经节不与alpha运动神经元直接联系,而是更好的保证运动执行。小脑与基底神经节有所不同,小脑可负责运动过程中的调节也可负责运动的发起,而基底神经节则负责运动发起。
八.联合皮质
联合皮质并不是为单一功能服务的脑区,它在感觉输入和运动输出之间起到联结的作用。联合皮质连接着感觉皮质与运动皮质,如果对这个脑区施加单独的刺激,无法观察到单一的反应。它的功能是对各种信息(即包括外源性视觉、听觉等信息,又包括内源性目的、动觉等信息)进行整合,然后将整合信息传入到运动皮层,从而调控运动。联合皮质约占人脑75%的区域,该比例远大于其他动物。
九.总结(Gazzaniga,lvry 和Mangun(著),周晓林和高定国等人(译),2011)
根据上述内容,可以将运动的神经系统总结如图20。
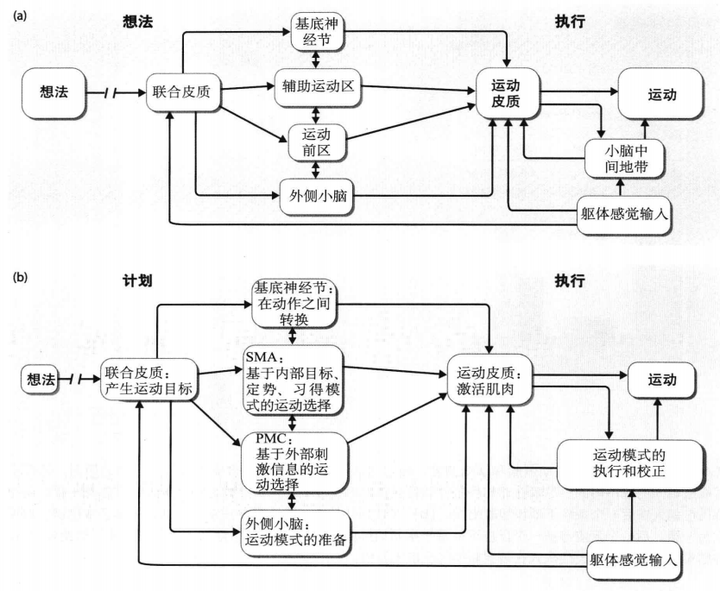
图20
十.神经元信息传递的知识基础(闫剑群(主编),2015)
在上面我们看到,同一个神经元会受到多个下行神经元控制。比如脊髓的神经元可能会受到初级运动区神经元和运动前区神经元的控制;一些神经元的传入信息可能是兴奋的,也可能是抑制的;另外,神经元的各种状态指的是什么,比如激活、抑制和易化等等。因此,我们需要对关于这部分的内容有一定的了解。
1.细胞结构
神经细胞包括胞体、轴突和树突。胞体是细胞的主体;每个细胞都有一个轴突,负责信息的输出,轴突上有多个轴突侧支,因此一个细胞可使信息传递到多个不同的细胞上;另外,每个细胞都有多个树突,负责接收不同细胞的传入信息。因为有些细胞的轴突可以长得很长,所以这些细胞可以与很远的细胞联系,当然它们也能与较近的细胞联系。
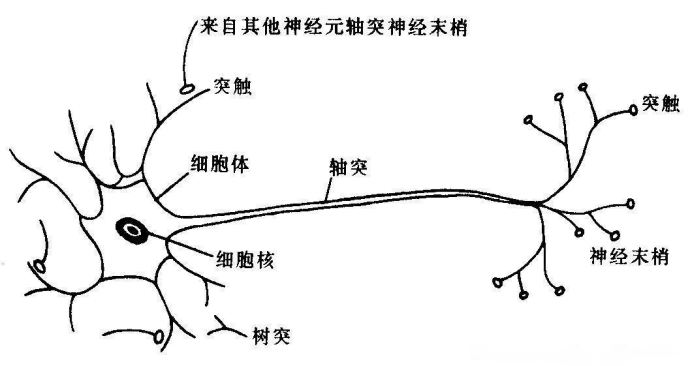
图21
轴突与其他细胞的连接处叫突触。突触的结构包括突触前膜,它是轴突发送信息的地方,而发送信号的细胞被称为突触前神经元;突触后膜,它是细胞接收信息的地方,而接收信号的细胞被称为突触后神经元;突触间隙,即突触前膜和突触后膜之间的间隙。突触前膜一般处于轴突处,突触后膜可以是树突、胞体和突触。因此突触的类型又可以分为轴突-树突式(图22(a))、轴突-胞体式(图(b))和轴突-轴突式(图(c))等。
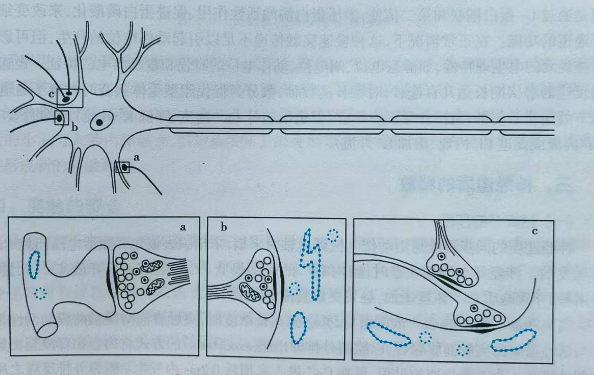
图22(a) 图22(b) 图22(c)
2.细胞的连接方式
细胞与细胞之间的连接方式有很多种,上面提及的脊髓细胞可以接收很多不同细胞的信息就是其中一种,即聚合式。细胞的连接方式包括:
(a)单线式联系:即一个突触前神经元仅与一个突触后神经元联系,例子有视锥细胞和神经节细胞的连接。如图23。
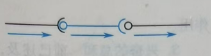
图23
(b)辐散式联系:一个突触前神经元通过轴突的不同分支与其他突触后神经元联系,从而对该信号进行广泛传播。多见于反射的传入通路。它能将同一个信息传递到不同神经元中。如图24。
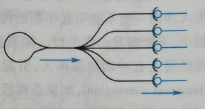
图24
(c)聚合式联系:刚刚提及的脊髓细胞就是聚合式。它指不同突触前神经元的突触汇聚于同一个突触后神经元,多见于反射的传出通路。它是神经元信息整合的基础。如图25。
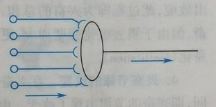
图25
(d)链锁式联系:即通过轴突的侧支与多个神经元联系,然后下一神经的轴突侧支又与下下个...。这种方式可以扩大神经元的影响范围。如图26。
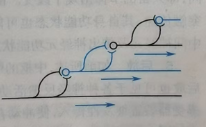
图26
(e)环状联系:指的是神经元在刺激一个中间神经元后,中间神经元又返回来对该神经元进行反馈。反馈可以是抑制性的,即负反馈,使刺激及时终止;也可以是兴奋性的,即正反馈,使刺激延续或增强。这种兴奋性环状联系可能是感觉记忆的基础。很多时候即使感觉刺激消失,我们仍然可以感受它,如视觉后效。如图27。
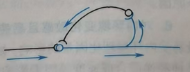
图27
(f)突触信息的整合
每个神经元都汇聚着大量其他神经元的输入,有些输入神经元是兴奋性的,有些则是抑制性的,有些输入信号强度高有些输入信号强度低,有些距离近有些距离远。最终突触后神经元会综合这些信息确定放电频率,这个过程被称为突触整合。
3.细胞的状态与活动
(1)静息电位与极化
一般情况下,细胞电位内部为负外部为正,并且只有较低的放电频率。这时该细胞所处的电位被称为静息电位,其状态又可被称为极化。极化是细胞的正常状态。
在极化状态下当细胞被传入兴奋信息时,细胞内电位变大及放电频率变大的过程叫做去极化。此时突触后神经元被激活,其电位被称为兴奋性突触后电位(EPSP);而在极化状态下当细胞被传入抑制信息时,细胞内电位得更少及放电频率被完全抑制,这个过程被称为超极化。此时突触后神经元被抑制,其电位被称为抑制性突触后电位(IPSP);如果电位从高位回落到静息电位,这个过程被称为复极化。
(2)动作电位与激活
动作电位指的是当去极化(兴奋)达到阈值时产生的电位。换句话说,当EPSP的总大小达到一个阈值时,突触后神经元会输出一个爆发性的电位,这个输出被称为动作电位。这时神经元的状态被称为激活。EPSP的总大小可以简化理解为各突触产生EPSP的总和,它包括两种形式的叠加:树突上不同部位EPSP的叠加和同一位置上在1-1.5ms内EPSP的叠加。实际上,这种叠加并非简单的数学意义的相加,每个突触输入对突触整合的影响与突触位置和重要性等因素相关。
(3)中枢抑制
中枢抑制可以分为突触后抑制和突触前抑制。
突触后抑制:当抑制性信息被传入时,突触后神经元上产生IPSP,产生超极化过程,使其处于抑制状态。有两种结构可以产生突触后抑制,分别对应传入侧支性抑制(图28(a))和返回性抑制(图28(b))。传入侧支性抑制是指,一个兴奋性信号传入一个抑制性中间神经元,使抑制性中间神经元对其下游神经元发送抑制信号并对其抑制。其中一个例子是原动肌和拮抗肌的配合。当兴奋性信号使原动肌激活收缩时,该兴奋性信号同样会激活一个中间神经元,使后者对拮抗肌产生抑制并使其放松。返回性抑制是指一个细胞在发出兴奋性信息时,通过激活一个抑制性中间神经元从而对自身产生负反馈。它的意义在于能及时终止神经元活动,并使同一中枢中的其他神经元同步活动。
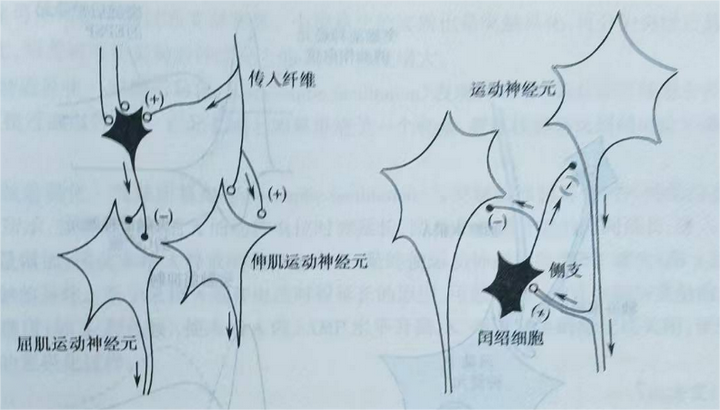
图28 (a) 图28(b)
突触前抑制:如图29,一个轴突A被另外一个轴突B介入(即轴突-轴突式),轴突B输兴奋性信息可使轴突A输出兴奋性信号的强度减少,最后使突触后神经元的EPSP明显减少,造成无法激活甚至不能引起兴奋。这种抑制对外周感觉的传入有重要意义,因为当特定感觉的神经元对其他感觉的神经元产生突触前抑制时,便可屏蔽其他感觉的干扰从而增强对该特定感觉的感知。
图29
(4)中枢易化
易化指的并非激活。它可以分为突触后易化和突触前易化。突触后易化指的是突触后神经元接收到兴奋信息,EPSP总大小接近到但还没到达动作电位阈值时的一种状态。这时只需再加上一个兴奋刺激就能使细胞激活。这就好比,突触后易化相当于把门锁打开,再给一个兴奋刺激相当于一个推门的动作,而门打开相当于激活。突触前易化的细胞结构与突触前抑制的相同,都是通过轴突-轴突式影响兴奋性信号传递。但不同的是,突触前抑制是使兴奋性信号强度减弱,而突触前易化是延长兴奋性信号的传入时间(最高强度不变),这相当于增加了EPSP时间上的总和,从而使突触后神经元的EPSP变大。
4.突触可塑性
这里有一种易化现象,当突触前神经元刺激突触后神经元产生放电现象后,下一次再次产生这种刺激时,放电电位的幅度要比之前的大。这种易化现象属于短期的突触塑造,它被认为是由于突触前神经元在放电之后还没达到静息电位,而新的刺激来临时促使更高强度的电位的发出。还有一种突触塑造叫做长时程增强,顾名思义这种塑造是长期的。它与学习、记忆有重要的关系。因此,突触并不是一成不变的,而是具有可塑性的。