本文内容速览:

在植物基因功能研究中,我们对"一个基因发挥什么作用"的解析路径已经相当熟悉。从突变体表型分析到过表达验证,再到表达模式、亚细胞定位以及分子互作等实验手段,围绕单个基因构建功能模型,已经形成了一套相对固定的研究范式(欢迎大家阅读我们之前出版的《植物基因功能研究范式》一书)。在大多数情况下,这样的研究路径足以支撑我们对一个基因功能的基本认知。然而,当我们试图在此基础上进一步解释其作用机制时,问题往往开始变得复杂。
在实际研究中,我们可以通过不同手段获得大量关于"上下游关系"的信息。例如,在转录调控层面,可以通过DAP-seq或ChIP-seq筛选转录因子的潜在结合位点,但体外检测到的结合位点数量往往远多于体内真实参与调控的位点(O′Malley et al., 2016);在表达层面,转录组数据中时常可以分析出一些基因表达具有高度相关性,但这种相关性也并不必然代表直接的调控关系(Kernfeld et al., 2008);在蛋白互作层面,酵母双杂或质谱筛选可以获得大量候选互作蛋白,但其中只有部分互作在体内具有稳定且有生物学意义的功能(Alvarez-Ponce et al., 2016);在蛋白修饰层面,大规模磷酸化组学可以检测到大量潜在修饰位点,但目前仅有一部分位点的功能已被解析,大多数仍需进一步的实验加以验证(Millar et al., 2019)。
这些结果提示我们,基因在细胞中并不是独立发挥作用,而是与其上下游共同发挥作用。然而,在不同层级的调控过程中,"能够检测到关联"与"真实发生调控"并不是同一个问题。也正是在这样的背景下,近年来植物分子生物学研究的一个趋势逐渐显现,即研究的重心正从"单基因功能解析"转向"调控网络解析"。与单基因研究不同,调控网络的核心并不在于连接更多的分子,而在于在复杂的关联关系中建立具有方向性和因果逻辑的调控链。因此,如何从多组学数据和多层级证据中区分"相关"与"因果"并合理验证这些潜在调控关系,逐渐成为调控网络研究中最关键的问题之一。
0 1

调控网络的构成

在明确了调控网络解析的核心问题之后,一个更具体的问题随之出现:对于一个具体基因而言,我们应该从哪里切入这一网络?在实际研究中,这个问题的答案常常并不清晰。很多情况下,研究路径更多的是由已有技术条件所决定,例如优先进行转录组筛选、互作蛋白筛选或某类组学分析,再围绕这些结果逐步补充上下游关系。然而,这种以技术为导向的切入方式,往往容易陷入一个共同的问题,即不断积累关联信息,却难以有效建立具有解释力的调控机制。从调控网络的角度来看,这一问题的本质并不在于选择哪种技术,而在于目标基因在整个调控网络中的位置。不同类型的基因,其在网络中的功能角色不同,对应需要优先回答的问题也不同。换句话说,研究的切入点并不取决于能做什么实验,而取决于这个基因在调控过程中扮演什么角色。
如果从信息流动的角度对调控网络进行拆解,可以将其大致理解为由多个功能层级构成。首先,信息感知层 ------一类分子负责感知外界或内源信号,其关键在于识别信号并发生状态变化;其次,信息处理层 ------部分分子处于信息传递和整合的中间环节,承担将上游信号向下游转导的功能;进一步向下,信息执行层 ------一些分子直接参与生理过程的执行,其功能最终体现在具体性状上;此外,信息调节层------还存在一类调控调节因子,通过影响关键节点的活性、稳定性或复合体状态,对整个网络进行精细调控。这些不同层级共同构成了调控网络的基本结构,那么这些层级包含哪些类型的基因,又如何研究呢,接下来给大家分享具体的文献案例。
1.1
信息感知层
从功能上来看,信息感知层主要是在接受信号的第一环节发挥作用,这一层通常包括几类典型分子。例如,位于细胞膜上的受体蛋白(如受体样激酶,receptor-like kinases,RLKs)可以感知激素或环境信号;部分感受器蛋白能够直接结合特定分子或离子;离子通道,例如钙离子通道,则通过改变胞内离子浓度实现快速信号转导;此外,一些与受体紧密关联的初级激酶,也可以在信号识别后迅速发生磷酸化状态变化,从而启动后续传导过程。从研究角度来看,以信息产生层为切入点,核心需要回答的问题通常是感知了什么信号?这一感知如何被转化为分子状态变化?这种变化如何向下游传递?
2024年5月,德国慕尼黑工业大学Erwin Grill课题组和Alexander Christmann课题组联合在Nature 杂志上发表了一篇题为"The temperature sensor TWA1 is required for thermotolerance in Arabidopsis "的研究论文,作者前期鉴定到了一个热胁迫响应相关基因TWA1 。通过突变体与互补实验证明TWA1 是拟南芥热耐受必需基因,进一步分析发现其位于热胁迫响应通路上游。关键的是,TWA1蛋白本身可对温度变化产生直接响应。基于这些结果,作者认为TWA1具有温度感受器的功能。在高温下,TWA1的构象发生改变,从而能够与JAM转录因子以及TPL和TPR蛋白发生物理相互作用,形成阻遏复合物,该复合体对热激转录因子HSFA2及下游热激蛋白的表达起正调控作用(图1),进一步对热胁迫做出反应。
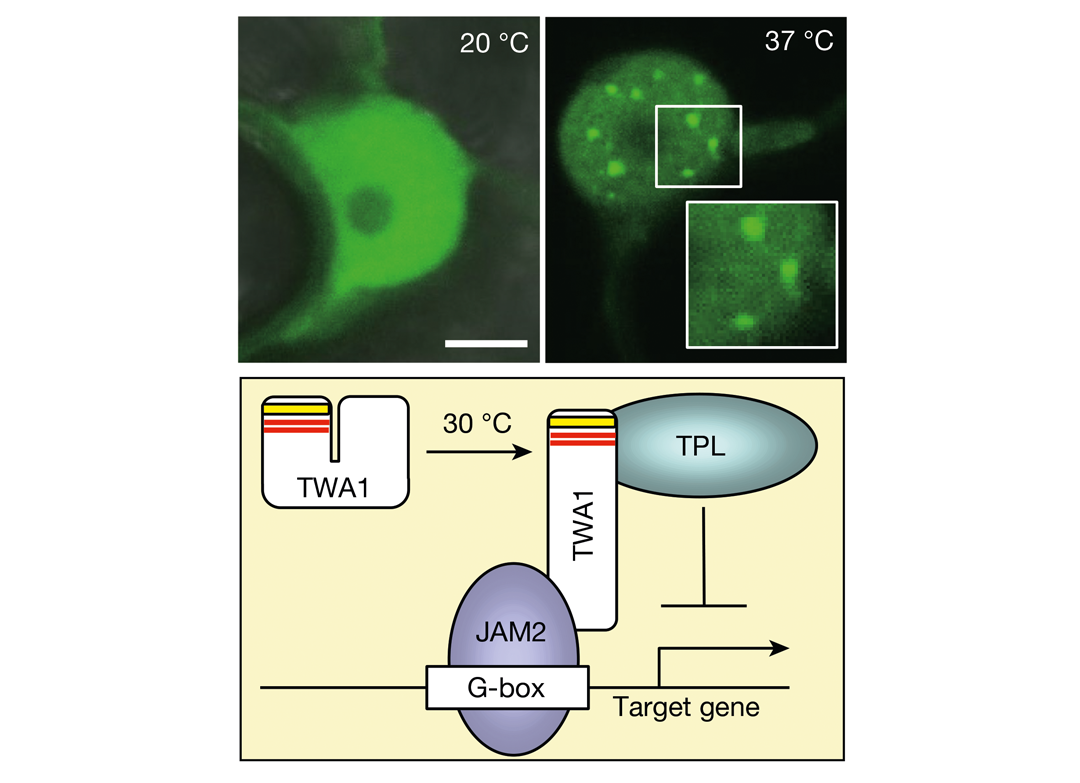
图1 高温前后TWA1的定位变化及作用模式图(Bohn et al., 2024)。
1.2
信息处理层
这一层主要负责对上游信号进行整合与传递,不同来源的信号在这一层汇聚。需要注意的是,这一层并不对应某一个固定节点,而往往由多个连续作用的调控组分构成,信号在不同节点之间依次传递、放大或整合。信息处理层通常包括转录因子(TF)、染色质调控因子以及RNA调控因子等。例如,转录因子通过识别特定顺式作用元件直接调控靶基因表达;染色质调控因子通过改变染色质结构或修饰状态影响基因的可及性;RNA调控因子则在转录后层面对基因表达进行精细调节。从研究角度来看,以信息处理层为切入点,核心需要回答的问题通常是:该分子如何接收上游信号?其调控的直接靶标是什么?在什么条件下发挥调控作用?
2026年1月,中国农业大学王宇课题组在New Phytologist杂志 上发表了一篇题为"ZmMPK5-mediated ZmOCL1 phosphorylation positively regulates drought tolerance by promoting the induction of ZmDHN2 in maize"的研究论文,该研究首先通过基因编辑突变体库筛选出了一个与干旱胁迫相关的转录因子ZmOCL1,进一步研究发现该转录因子能调控下游干旱胁迫直接相关基因ZmDHN2(脱水蛋白基因)的表达。此外,作者发现ZmOCL1介导的转录调控受到上游激酶ZmMPK5的磷酸化调控(图2)。最终,作者构建了一条参与调控玉米耐旱的ZmMPK5-ZmOCL1-ZmDHN2调控通路。
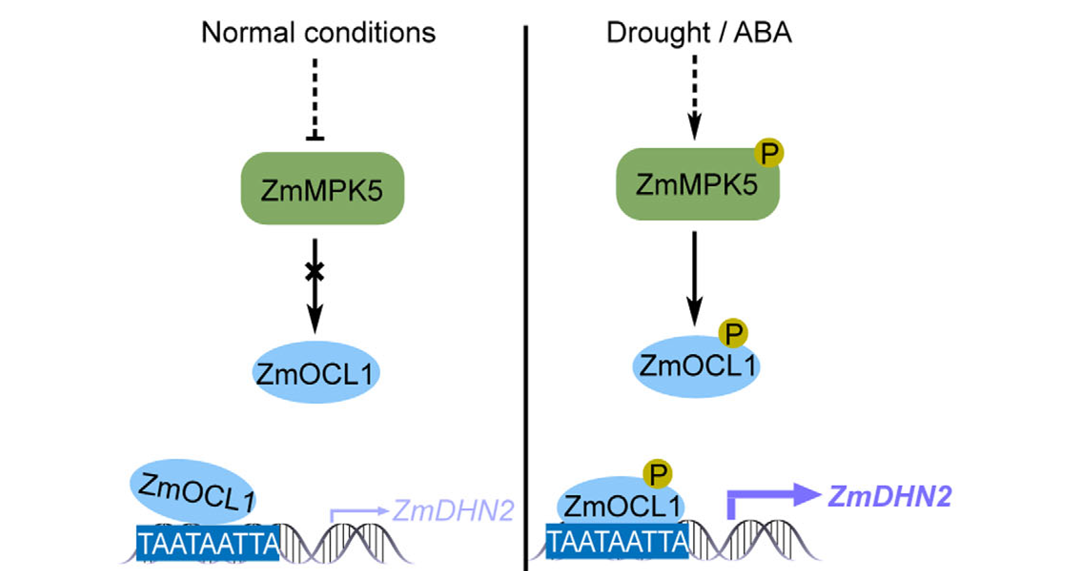
图2 ZmOCL1在玉米干旱响应调控中的工作模型(Yang et al., 2025)。
1.3
信息执行层
从功能上来看,这一层主要负责将上游信号转化为与表型直接相关的生理过程,是直接影响植物表型的部分。信息执行层层通常包括代谢酶、转运蛋白以及结构蛋白等。例如,代谢酶通过调控代谢通路影响代谢产物的积累;转运蛋白通过调节物质跨膜运输改变细胞内环境;结构蛋白则参与细胞结构或组织形态的构建。从研究角度来看,以信息执行层为切入点,核心需要回答的问题通常是:该分子具体参与了哪些生理过程?其功能如何在表型层面体现?以及这一功能受到哪些上游调控因子的控制?
2026年4月,湖南农业大学刘仲华、刘硕谦与田娜课题组在Plant Biotechnology Journal 杂志上发表了一篇题为"Transcriptional regulation of the novel theacrine synthase gene CsTcS2 by the CsTINY --CsWRKY33 module underpins theacrine biosynthesis in Camellia sinensis "的研究论文,该研究首先从高苦茶碱茶树中鉴定到了一个新的苦茶碱合成酶基因CsTcS2 ,并通过转基因功能验证以及酶催化能力检测证明了其在茶碱生物合成途径中发挥着催化酶的作用。为了进一步研究CsTcS2 受到上游哪些转录因子的转录调控,作者鉴定分析到一个转录因子CsTINY可以正向调控CsTcS2 的表达。此外,作者还发现转录因子CsWRKY33 的表达也受到CsTINY的调控,同时CsWRKY33还能与CsTINY互作协同调控CsTcS2 的表达,这进一步完善了CsTcS2的转录调控通路(图3)。
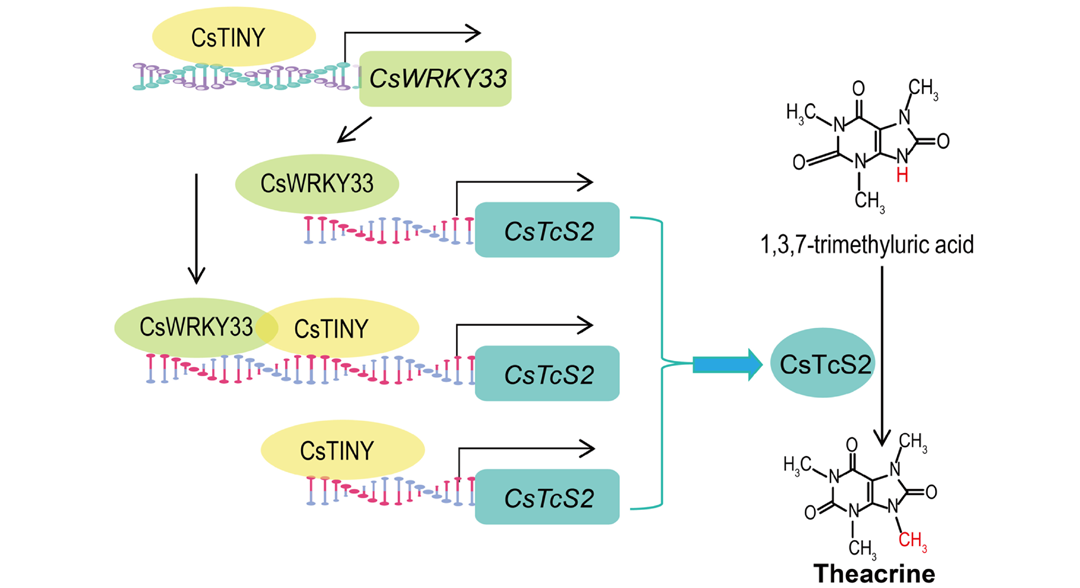
图3 CsTINY-CsWRKY33介导的CsTcS2表达和茶碱生物合成调控的工作模型(Wu et al., 2026)。
1.4
信号调节层
从功能上来看,这一层并不直接参与信号的传递以及直接影响表型相关的生理过程,而是通过调节其他几个层级的基因从而对整个调控网络进行精细控制。信息调节层通过改变蛋白的活性、稳定性、定位或复合体状态,从而影响信号传递或执行过程的强度与持续时间。该层通常包括E3泛素连接酶、蛋白修饰酶及分子伴侣等。例如,E3泛素连接酶可以通过调控靶蛋白的降解改变其丰度;蛋白修饰酶可以通过磷酸化或其他修饰影响蛋白活性;分子伴侣可以通过影响蛋白折叠与稳定性参与调节过程。以调控调节层为切入点,核心需要回答的问题通常是:该分子调节了哪些关键节点?通过何种方式影响其功能?这种调节在什么条件下发生,并如何改变整个调控通路的输出?
2025年6月,中国农大张永亮课题组与加州大学戴维斯分校Savithramma P. Dinesh-Kumar课题组合作在Science Advances杂志上发表了一篇题为"A phosphorylation-driven ubiquitination switch fine-tunes Alfin-like 7-induced ROS signaling in plant immunity"的研究论文,该研究聚焦于植物免疫过程中的ROS稳态,深入研究了植物细胞如何通过精细调控ROS的积累和消除,从而平衡植物的生长与防御。作者在前期研究中发现,转录抑制因子AL7通过抑制ROS清除基因的表达来促进ROS积累,从而正调控核苷酸结合富亮氨酸重复(NLR,Nucleotide-Binding Leucine-Rich Repeat)受体介导的植物免疫,并且这一过程受丝裂原活化蛋白激酶(MAPK,Mitogen-Activated Protein Kinase)介导的Ser174位点磷酸化的正向调控(Zhang et al., 2023)。该文章进一步研究发现,在前期明确的调控通路中,AL7还受到了泛素化的调控。AL7在Ser174位点的磷酸化增强了其与E3泛素连接酶UBR7的相互作用,从而增加了AL7的泛素化水平并促进了AL7的降解,这种降解恢复了ROS清除基因的表达,防止ROS的过度积累(图4)。
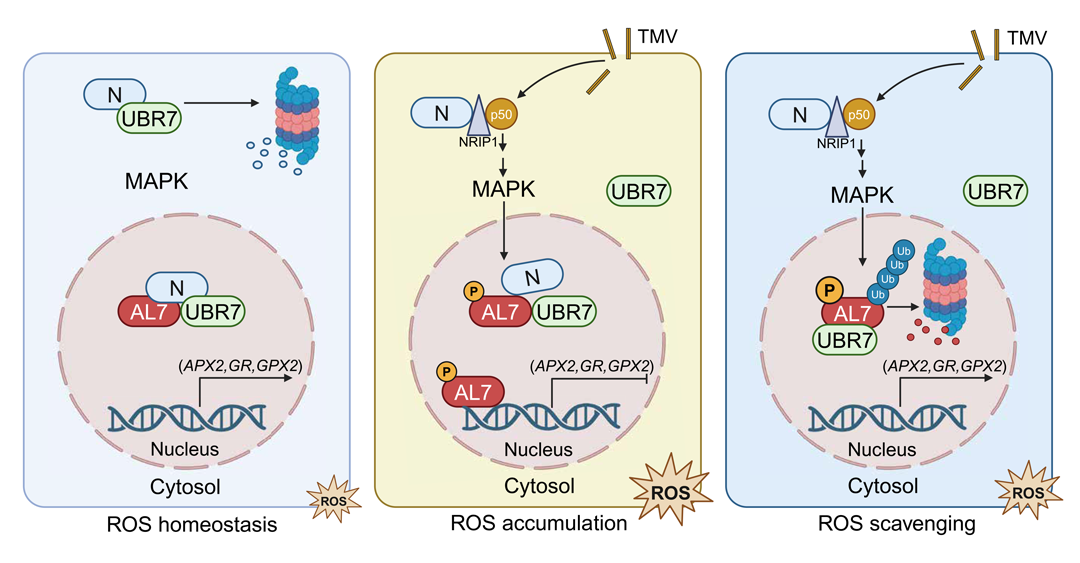
图4 磷酸化驱动的泛素化开关模型,该开关可微调NLR介导的免疫反应期间AL7诱导的ROS信号传导(Zhang et al., 2025)。
02

解析 调控网络 的共同原则

通过前面的案例可以看到,调控网络的解析,本质上是以目标蛋白为锚点,在判断其在表型调控过程中的功能层级之后,逐步分析其与上游调控因子及下游作用对象之间的关系,并通过实验加以验证。在这一过程中,虽然不同层级的切入方式有所差异,但在具体分析逻辑上,仍然呈现相对一致的特点。
1)明确目标基因在调控网络中的位置
调控网络的解析通常始于对目标基因功能角色的判断,而不是实验手段的选择。不同层级对应的问题本身存在差异,如果在未明确其所处位置的情况下直接开展分析,往往容易导致研究偏离核心问题。因此,在进入具体实验之前,对其在调控通路中的大致定位进行判断,是后续研究能够有效展开的前提。
2)区分相关性与因果关系
在不同层级的研究中,表达变化、组学关联以及蛋白互作等信息都可以作为分析线索,但这些结果本身并不直接等同于调控关系。相关性可以用于提出假设,但只有在明确作用方向并获得直接证据支持之后,相关关系才有可能被归入调控通路的框架之中。
3)多层级证据的相互支撑
调控关系的建立通常依赖不同类型证据的结合。组学分析有助于筛选潜在的上下游关系,分子实验用于验证其直接作用方式,而遗传学证据则从整体水平上评估其生物学意义。不同层级证据之间相互补充,使得调控关系能够从"可能存在"逐步成为"可以成立"。
4)回归性状层面的解释
调控网络的构建并非目的本身,其意义在于解释具体的生物学过程或表型变化。如果调控关系无法与实际性状建立联系,会让人感觉研究没有"落地"。因此,无论从哪个层级切入分析,最终都需要将分子层面的关系整合到性状层面进行理解。
在明确了上述基本原则之后,还需要进一步考虑一个更实际的问题:调控网络究竟需要解析到什么程度。从目前的研究来看,不同文章的深度存在较大差异,不同期刊的侧重点也并不相同,因此很难给出一个统一的标准。对于具体研究而言,更重要的是围绕目标蛋白这一核心,说明其在调控网络中的位置,以及其如何参与调控并受到调控,而不是对所有关联蛋白都进行全面而细致地分析。换句话说,调控网络的解析的精髓不在于面面俱到,而在于关键的调控关系是否被解析清楚。
小远叨叨


近年来,国内植物领域的高水平文章频发,就算是CNS这些顶刊也常见有研究成果出现。纵观这些研究可以发现,相较于早期以单基因功能解析为主的研究范式,当前工作正明显向关键科学问题驱动的分子调控网络解析转变,研究深度与系统性都在提升。随着多种组学还有各种验证手段的进步以及AI领域的发展,使得解析复杂分子调控网络具备了一定的先决条件。然而,在实际研究过程中,如何使用各种技术手段解析海量数据,合理筛选关键节点并完成因果关系验证,仍然是当前调控网络研究中的核心挑战之一。本次推文以调控网络如何解析的角度出发,希望可以在具体研究中给大家提供借鉴,大家在课题研究过程中如果有任何问题,非常欢迎随时联系伯远生物!
References:
Alvarez-Ponce D. Recording negative results of protein--protein interaction assays: an easy way to deal with the biases and errors of interactomic data setsJ. Briefings in Bioinformatics, 2017, 18(6): 1017-1020.
Bohn L, Huang J, Weidig S, et al. The temperature sensor TWA1 is required for thermotolerance in Arabidopsis J. Nature, 2024, 629(8014): 1126-1132.
Kernfeld E, Keener R, Cahan P, et al. Transcriptome data are insufficient to control false discoveries in regulatory network inferenceJ. Cell systems, 2024, 15(8): 709-724. e13.
Millar A H, Heazlewood J L, Giglione C, et al. The scope, functions, and dynamics of posttranslational protein modificationsJ.Annual review of plant biology, 2019, 70(1): 119-151.
O'Malley R C, Huang S C, Song L, et al. Cistrome and epicistrome features shape the regulatory DNA landscapeJ. Cell, 2016, 165(5): 1280-1292.
Wu T, Zhu L, Shao C, et al. Transcriptional Regulation of the Novel Theacrine Synthase Gene CsTcS2 by the CsTINY--CsWRKY33 Module Underpins Theacrine Biosynthesis in Camellia sinensis J. Plant Biotechnology Journal, 2026.
Yang X, Cheng J, Wang F, et al. ZmMPK5‐mediated ZmOCL1 phosphorylation positively regulates drought tolerance by promoting the induction of ZmDHN2 in maizeJ. New Phytologist, 2026.
Zhang D, Gao Z, Zhang H, et al. The MAPK-Alfin-like 7 module negatively regulates ROS scavenging genes to promote NLR-mediated immunityJ. Proceedings of the National Academy of Sciences, 2023, 120(3): e2214750120.
Zhang D, Zhang X, Wen Z, et al. A phosphorylation-driven ubiquitination switch fine-tunes Alfin-like 7--induced ROS signaling in plant immunityJ. Science Advances, 2025, 11(24): eadw7554.