本文内容速览:
遗传转化是作物改良、基因功能研究和分子育种的关键工具。然而,传统方法受限于基因型依赖性、操作复杂以及转化效率低等问题,导致许多物种的研究受到了限制。在先前的推文"植物高效遗传转化方法,请查收!"中,小远给大家介绍了GiFT、RAPID、RBS、ES-CDB以及SAMT这几种快速、高效的遗传转化方法,它们在提高转化效率的同时,不仅在很大程度上克服了基因型依赖性,还极大地简化了转化流程。科研工作者开发和优化遗传转化方法的脚步从未停歇,时隔将近两年,小远又收集了不少新的资料,在本期推文中将继续跟大家分享,接下来就跟着小远一起来看看都有哪些具体的内容吧。
ROTIS
繁琐的组织培养、漫长的转化周期以及基因型依赖性等因素极大地限制了大豆分子育种和基因功能研究的进程。因此,开发出操作简便、高效且普适的大豆遗传转化体系是该领域亟待解决的问题。
2025年12月,东北农业大学韩英鹏课题组联合中国农业科学院作物科学研究所/国家南繁研究院和玉兵课题组在Plant Physiology 杂志上发表了一篇题为"ROTIS: A RUBY-assisted one-shot tissue-culture-free system for in-planta soybean transformation from germinated seedlings"的研究论文,该论文报道了一种名为ROTIS(RUBY-assisted One-shot Tissue-culture-free In-planta Soybean-transformation)的遗传转化系统,该系统以萌发大豆为外植体,将非组培条件下的农杆菌转化与RUBY红色可视化筛选标记相结合,成功构建了一套高效、易操作的大豆转化方案。
作者首先将大豆播种在珍珠岩中,在黑暗条件下用0.1mg/L的6-苄氨基嘌呤(6-BA)溶液灌溉,使其萌发。待子叶在弱光条件下完全展开并变为绿色后,去除种皮并轻轻分离子叶使顶端分生组织暴露出来。使用无菌手术刀切除顶端分生组织,在生长点处制造伤口。将处理后的幼苗外植体浸入含有EHA105农杆菌的共培养培养基悬浮液中进行真空渗透处理,然后用浸有农杆菌的棉球继续处理伤口,以确保农杆菌可以侵染到外植体中。在22℃、低光照条件下进行48小时的共培养,随后转入标准光照条件,并每隔48小时更换一次棉球。共培养3-8天后,通过定期更换浸有芽诱导培养基和卡那霉素的棉球进行不定芽的诱导和筛选。通常在农杆菌侵染后的14-21天,抗性不定芽簇会从伤口处萌发,此时可以停止使用芽诱导培养基。待不定芽伸长且第一片三出复叶完全展开后,使用卡那霉素(500mg/L)溶液涂抹叶片进行筛选,将无抗性的再生芽从基部切除。连续三轮三出复叶筛选中表现出持续抗性的幼苗,随后可以进行转基因阳性检测(图1)。整个流程仅需40-80天即可获得转基因植株,极大地缩短了获得转基因大豆的周期。
为了进一步简化筛选过程并提高效率,作者在表达载体中引入了由CaMV 35S启动子驱动的RUBY报告基因(图1B)。在共培养后的3-5天内,农杆菌侵染成功的部位会逐渐出现红色色素沉着,通过肉眼即可识别。在芽诱导过程中,新形成的不定芽在萌发时就会呈现红色,或在发育过程中逐渐积累色素,当伸长的不定芽叶片上呈现出清晰均匀的红色时,即可将整株幼苗移植到土壤中。RUBY报告基因的引入使ROTIS系统真正实现了转化的"可视化"。

图1 大豆转化流程及RUBY转基因株系在不同发育阶段的表型特征(Li et al., 2025)。(A1)大豆外植体状态;(A2、A3)切除茎尖主芽;(A4)在含有农杆菌的共培养培养基悬浮液中对外植体进行真空渗透处理;(A5)将含有农杆菌悬浮液的棉球放置于外植体上;(A6)用夹子固定外植体的两片子叶;(A7、A8)大豆丛生芽诱导过程;(A9-A11)大豆不定芽的再生与筛选;(A12)移除未转化芽并进行过渡培养;(A13)抗性芽的筛选与培养:通过抗性筛选获得表现出转基因抗性的再生芽;(B)RUBY表达载体的T-DNA区域示意图;(C1)与农杆菌共培养后,RUBY植株在子叶节处呈现出红色表型;(C2-C5)RUBY阳性芽的再生过程;(D1、D2)RUBY阳性叶片的表型;(E1-E3)RUBY阳性幼苗的花芽表型;(F1-F6)RUBY转基因阳性幼苗的果荚和种子表型;(G)RUBY转基因大豆深红色种子萌发后的表型;(H)大豆遗传转化流程示意图;(I)PCR检测RUBY基因的结果;(J1、J2)RUBY植株叶片红色深浅的表型差异及相应的GUS组织化学染色结果;(K1、K2)RUBY大豆花朵中的红色色素沉着及相应的GUS组织化学染色结果;(L1、L2)RUBY大豆荚果中的红色色素沉着及相应的GUS组织化学染色结果;(M1、M2)RUBY大豆茎部的红色色素沉着及相应的GUS组织化学染色结果;(N、O)对红色深浅程度不同的大豆叶片中Ruby和Gus基因进行RT-qPCR分析。
为了进一步验证ROTIS系统的普适性,作者选取了21个不同地理区域的大豆栽培品种进行测试。农杆菌侵染5-8天后,28.6%至64.3%的外植体出现红色的表型,经筛选和诱导不定芽再生后,获得了不同基因型大豆的RUBY转基因阳性株系。对转基因大豆植株进行PCR检测和表型分离比分析表明,外源基因能够稳定遗传,后代符合孟德尔遗传定律(图2)。这些结果证实了ROTIS系统具有广泛的基因型独立性,适用于多种基因型的大豆。
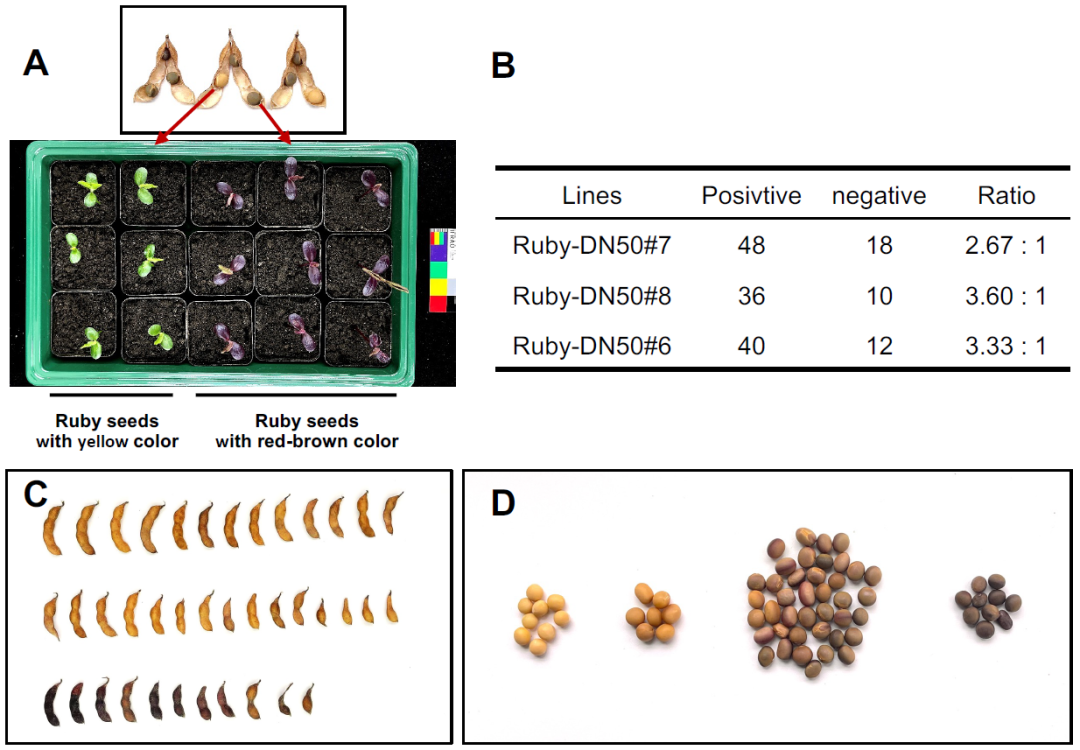
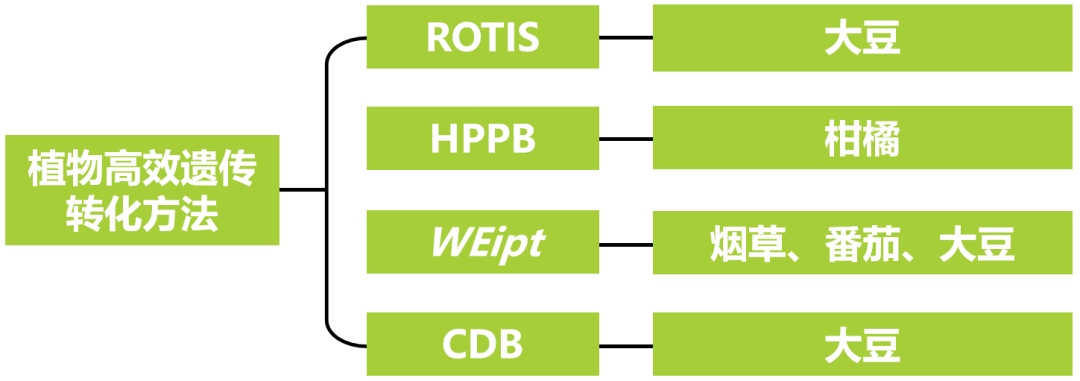
图2 T0代RUBY转基因株系后代的分离情况及表型(Li et al., 2025)。(A)从T0代收获的黄色Ruby大豆种子播种后,萌发出的幼苗呈绿色,表明这些是分离后不含RUBY基因的T1代大豆;从T0代收获的深红褐色Ruby大豆种子播种后,萌发出的幼苗呈RUBY特有的深紫红色,表明这些是分离后含有RUBY基因的T1代大豆,即转基因阳性大豆;(B)RUBY转化大豆株系T1代植株的分离比;(C、D)T0代RUBY转基因大豆成熟豆荚和种子的表型特征。
HPPB
柑橘砧木的根系不仅对水分和养分的吸收至关重要,还有助于与土壤细菌建立有益的联系(Song et al., 2023)。利用遗传转化技术来增强砧木的抗逆性、抗病性以及土壤微生物互作能力,不仅能有效的提升接穗的果实品质,还可以减少农药的使用(D'Amico et al., 2018; Zhang et al., 2022)。因此,建立一种高效、稳定且无需组织培养的柑橘砧木遗传转化技术,将为抗病育种、抗性评价及其田间落地应用提供强有力的技术支持。
2025年4月,赣南师范大学段硕课题组联合李敏课题组在Plant Biotechnology Journal杂志上发表了一篇题为"Establishment and application of high-pressure propagation breeding (HPPB)-mediated genetic transformation system in citrus rootstocks"的研究论文,该论文报道了一种基于高压繁育(High-Pressure Propagation Breeding,HPPB)的高效柑橘砧木遗传转化系统,通过将发根农杆菌转化与高压繁殖盒相结合,在田间环境下就可以快速获得遗传转化根系。
作者首先将含有目的基因载体的K599农杆菌在YEP培养基中,培养至OD600达到0.6-0.8的范围。接着,收集菌株并在MES溶液中进行重悬,在黑暗条件下培养2-4小时来激活农杆菌诱导生根的功能。选择2-3年生的柑橘作为外植体,去除茎上的刺和枝条后,用刀片在茎上切割出深度足以露出韧皮部并到达木质部的切口,将浸有K599重悬液的吸水纸贴在伤口处20分钟。最后,用预先填充好基质的灰色哑光高压繁育盒覆盖在伤口区域,并向盒中注射1-2mL农杆菌重悬液,在温室条件下培养。转化两周内,转基因根就会开始萌发,继续培养1-2个月即可获得大量转基因根系。随后,可通过WB和RT-qPCR分析转基因根中的基因表达情况,经过验证的转基因根可以被切下并移植,从而建立新的砧木植株(图3)。在表达载体中引入GFP报告基因后,利用手持紫外荧光灯就可以进行初步筛选,也可以利用RUBY,通过肉眼观察红色表型来快速筛选,总之,利用GFP或RUBY标签可以实现在田间环境下进行快速初筛(图4)。在HPPB过程中,对砧木进行嫁接来引入系统性病害,可以同时获得转基因根和病害感染根系,从而为致病相关机制研究提供研究模型。此外,作者还在HPPB中验证了基因编辑的效率,确定了其可行性。

图3 HPPB介导的遗传转化:从操作流程到实际应用(Zhang et al., 2025)。(a、b)第一部分HPPB介导的遗传转化示意图:(a)获取经病原菌侵染的转基因根系材料,(b)获取转基因根系材料;(c-e)第二部分HPPB在不同砧木中的应用;(f-h)第三部分HPPB在CRISPR基因编辑中的应用;(i)第四部分利用HPPB方法转化不同基因。

图4 GFP和RUBY标签的应用效果(Zhang et al., 2025)。
WEipt
建立对组织培养依赖度低、可直接实现植株原位(in planta)再生的技术体系,是当前许多植物遗传转化与基因编辑亟待突破的重要研究课题。
2025年11月,得克萨斯理工大学Gunvant B. Patil课题组在Molecular Plan t 杂志上发表了一篇题为"A synthetic transcription cascade enables direct in planta shoot regeneration for transgenesis and gene editing in multiple plants"的研究论文,该论文报道了一种创新的方法,利用WIND1 -ESR1级联反应来加速芽从头再生,在幼龄烟草和番茄的节间以及大豆胚轴(缺乏分生组织活性)部位,直接诱导产生了可遗传的转基因或基因编辑新芽。该技术有望在不依赖或仅极少组织培养的情况下,加速转基因和基因编辑植株的培育。
植物受伤后会激活一系列基因来修复组织。WIND1作为创伤诱导细胞重编程的核心调节因子,被激活后会直接结合到ESR1 启动子中与维管组织及创伤响应相关的顺式作用元件上,启动ESR1 的表达(Iwase et al., 2017)。作者以此为基础设计了一系列级联表达载体,在烟草中验证发现转化pro35S::WIND1 -proESR1 ::RUBY 载体的外植体在无植物激素的MS培养基上形成的愈伤组织与对照组相比增加了3-4倍。证实级联系统有效后,作者将proESR1 下游的RUBY 替换为ipt 基因,在烟草外植体中进行了测试,结果显示pro35S::WIND1 -proESR1 ::ipt (WEipt )载体显著增强了愈伤组织形成,并在无植物激素的培养基上诱导了多个芽分生组织的形成。这一结果证实,由AtWIND1 介导的proAtESR1 驱动ipt的表达可有效诱导芽从头再生(图5)。
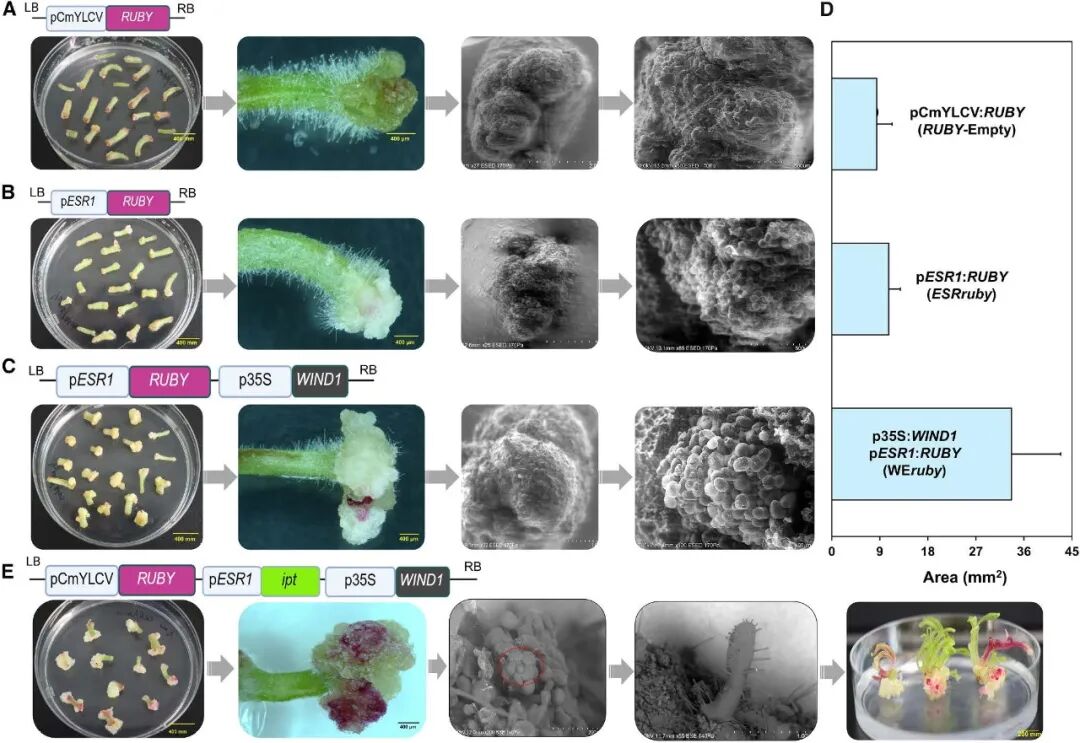
图5 体外验证"合成级联"的可行性(Kshetry et al., 2025)。(A-C)携带CmYLCV::RUBY (A)或proAtESR1 ::RUBY (B)载体的根癌农杆菌(GV3101)转化烟草茎段外植体后,仅形成少量愈伤组织或无愈伤组织;而当共转化35S::AtWIND1 -proAtESR1 ::RUBY 时(C),愈伤组织形成量显著增多,同时AtESR1 启动子被明显激活;(D)愈伤组织面积的定量分析显示,转化CaMV 35S::AtWIND1 的外植体,其愈伤组织形成量约为对照的3-4倍;(E)AtWIND1激活AtESR1 启动子驱动DR基因(如ipt)的表达,可在无外源植物激素的培养基中快速诱导愈伤组织产生,并促使茎尖分生组织形成,进而诱导新生分生组织的发生。
接着,作者将多个已报道的DR基因,包括ZmWUS 、AtSTM 、ipt 、GmBBM 和GmGRF -GIF 克隆至proESR1 下游构建载体,载体中还包含由拟南芥U6-snRNA26(U6-26)启动子驱动的靶向烟草PDS 基因的gRNA;由CaMV 35S启动子驱动的AtWIND1 以及由CmYLCV 启动子驱动的可视化报告基因RUBY (图6A)。利用农杆菌转化法将这些载体转入表达Cas9 的转基因烟草植株中,结果显示,发现将含ipt 的载体注射到剪去顶端的幼年烟草的茎切面时,效果最佳(图6B),可实现71%的芽再生效率,转基因与编辑效率分别为35%、10%。其它DR基因,除了GmGRF -GIF可在注射位点诱导出极少的芽,剩余的均未能诱导再生新芽。
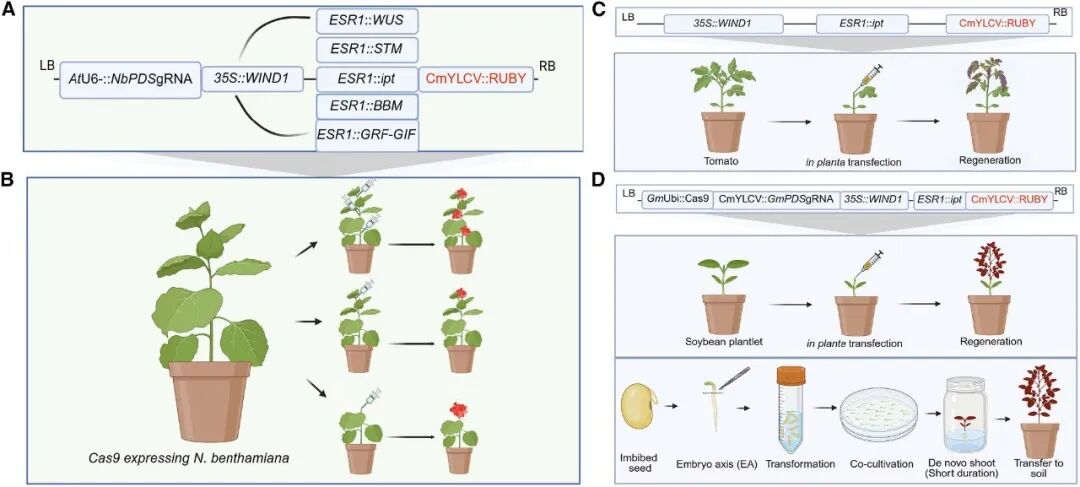
图6 表达载体与转化策略示意图(Kshetry et al., 2025)。(A)表达载体示意图;(B)烟草中的三种不同植物体内转化策略,分别在多个节间、靠近茎端分生组织的节间区域、剪去顶端后的茎段切面处进行转化;(C)番茄中表达载体及植物体内转化与再生策略的示意图;(D)大豆中表达载体及转化与再生策略的示意图:上方为植物体内转化策略(诱导了愈伤组织形成,但未能成功再生新芽),下方为半植物体内转化策略。
最后,作者将WEipt 转化体系应用到番茄和大豆中来评估其普适性(图6C、D)。在去除顶端的番茄幼苗上进行本体注射后,成功诱导出了再生新芽,再生效率约52%,转基因效率约21%。不过,遗憾的是尽管该方法成功再生出了表达大量甜菜红素的芽,但这些转基因芽无法形成可育果实(图7D)。这可能是因为甜菜红素的高积累或WEipt载体在番茄中的物种特异性效应导致的。在大豆中,直接采用茎秆注射的原位转化方法,无法有效诱导再生新芽。作者转而使用大豆胚轴作为外植体,在无菌条件下进行农杆菌转化和短暂共培养后,便将其转移到不含任何激素的培养基上,最终成功诱导出了新芽,再生效率可达80%,转基因效率为28%,基因编辑效率超12%,各项指标均显著优于传统大豆组织培养技术(图8)。
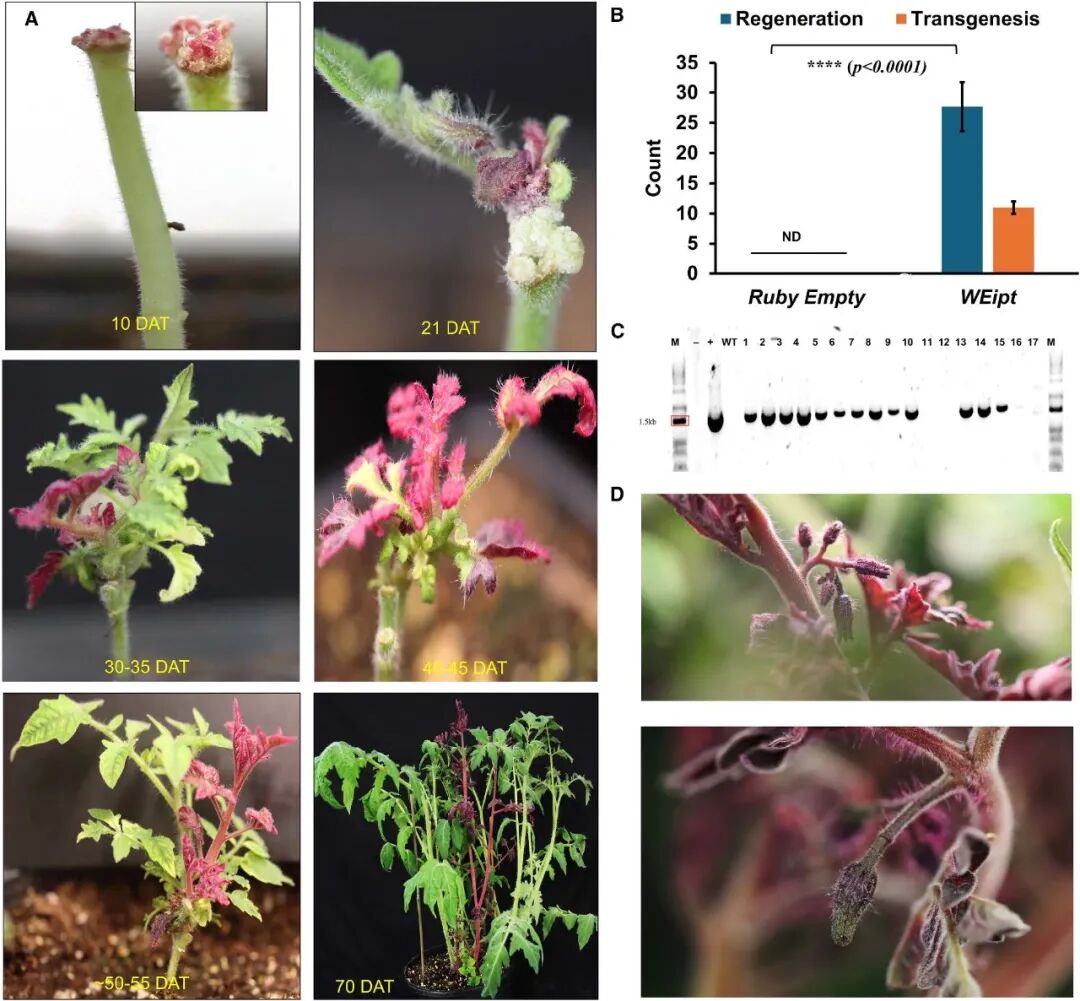
图7 番茄的植株原位转化与芽从头再生(Kshetry et al., 2025)。(A)番茄芽从头再生的各个阶段,展示了经WEipt载体转化后,从伤口部位长出的代表性转基因芽(红色组织);(B)再生芽中转基因数量的定量分析,反映了转化效率;(C)凝胶电泳图,通过PCR分析T0代是否为转基因植株;(D)积累了高水平甜菜红素的转基因芽所开的花。
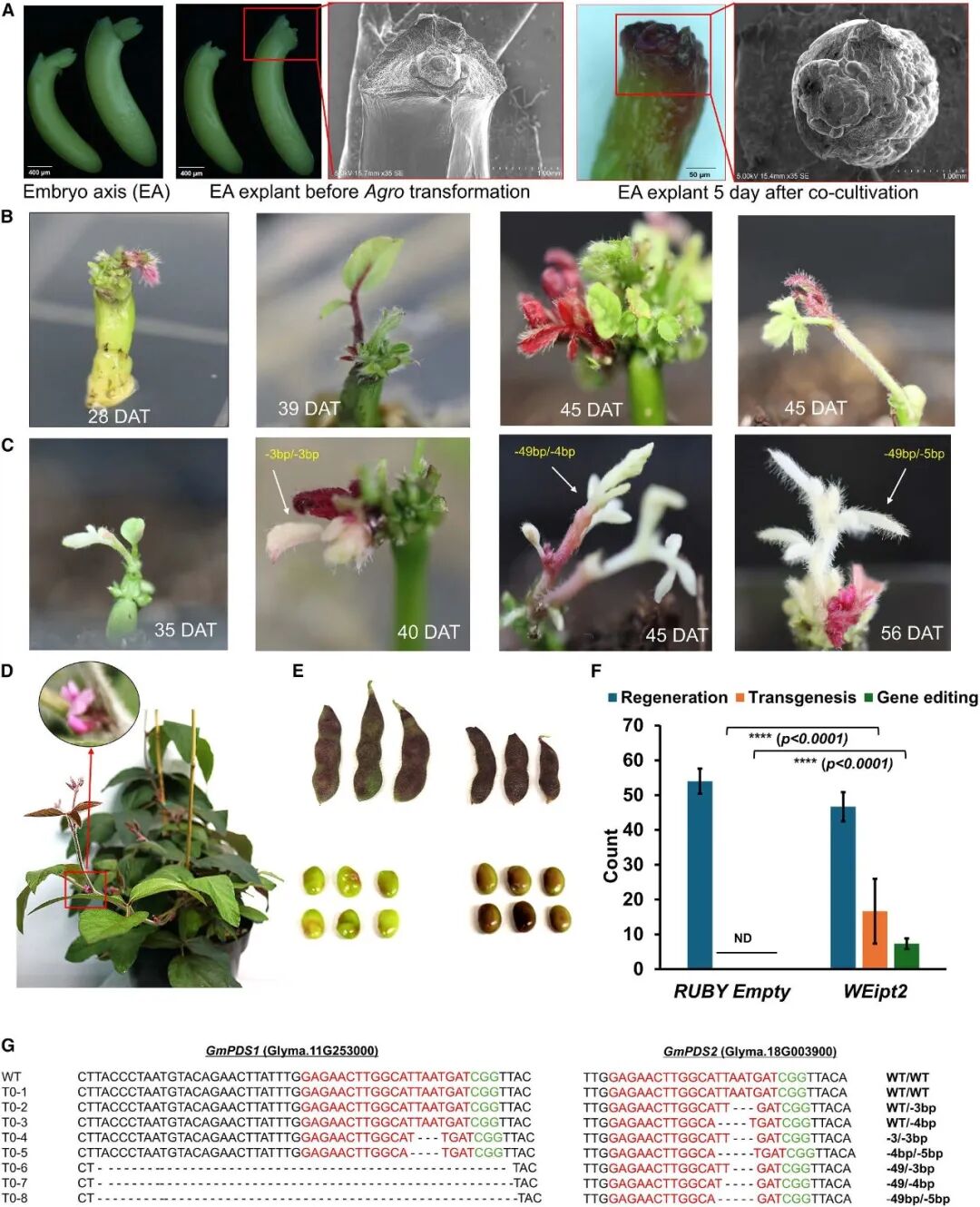
图8 大豆半无组织培养转化体系(Kshetry et al., 2025)。(A)用于大豆转化的胚轴(EA)外植体图像,扫描电镜图像显示了转化3天后愈伤组织的形成情况;(B)转化WEipt2 载体的胚轴外植体中,芽从头再生的不同阶段及RUBY 基因的表达情况;(C)转化WEipt2 载体的胚轴外植体再生的代表性基因编辑芽(半白化/白化芽);(D)转化后40-45天,将转基因芽移栽至土壤中,继续生长至成熟;(E)转基因植株的果荚和种子;(F)再生芽中的转基因和基因编辑统计数据;(G)T0代编辑芽中GmPDS1 和GmPDS2的编辑情况分析。
CDB
CDB(Cut-Dip-Budding)技术是基于多种植物离体组织的再生潜力开发的,通过简单的切割,蘸取农杆菌侵染,直接在植物切口处诱导转基因芽形成。在先前的推文中小远有给大家简单介绍过该技术在橡胶草中的应用,本次推文则主要给大家介绍该体系在大豆中的应用。
2025年12月,南方科技大学朱健康课题组在The innovation杂志上发表了一篇题为"An efficient tissue-culture-free soybean genetic transformation technology using the extremely simple cut-dip-budding strategy"的研究论文,将CDB策略成功拓展至大豆中,建立了一套无需组织培养,不依赖基因型且操作简单的大豆遗传转化及基因编辑体系。
在该论文中,作者首先评估了多种类型的大豆外植体的再生能力。结果显示,子叶腋区损伤或初生叶被切除的幼苗,以及子叶腋区损伤或切除胚芽的吸胀种子均能形成再生芽(图9A)。考虑到外植体制备的易操作性,作者选择去除胚芽的吸胀种子作为外植体来开发大豆CDB转化体系。
首先,将大豆种子用水浸泡12-16小时后,去除种皮和一片子叶,并切除胚芽。接着,将含有目的载体的农杆菌菌体用侵染缓冲液(含10mM MgCl2和10mM MES,pH 5.4)重悬。准备完毕后,将去除胚芽的吸胀种子浸泡在农杆菌悬浮液中先进行真空渗透15分钟,然后震荡孵育1小时,之后将外植体插入土壤中培养。培养期间,通过喷洒20mg/L的壮观霉素溶液来筛选转基因芽,并定期将顶芽和非转基因芽切除,仅保留转基因阳性芽(图9B-D)。按照上述操作流程最终获得阳性再生芽的效率为0.42%,为了提高效率,作者除了将发育调节因子引入表达载体中,还测试了细胞分裂素处理,结果显示,引入ipt 效果最佳(9E)。将ipt与其它发育调节因子组合使用可以进一步提高再生和转化效率(9G)。
作者在多个大豆品种中测试CDB转化体系均有效果,这表明该体系不依赖基因型,具有普适性。并且,通过CDB转化体系获得的转基因或基因编辑幼苗具备遗传转稳定性。
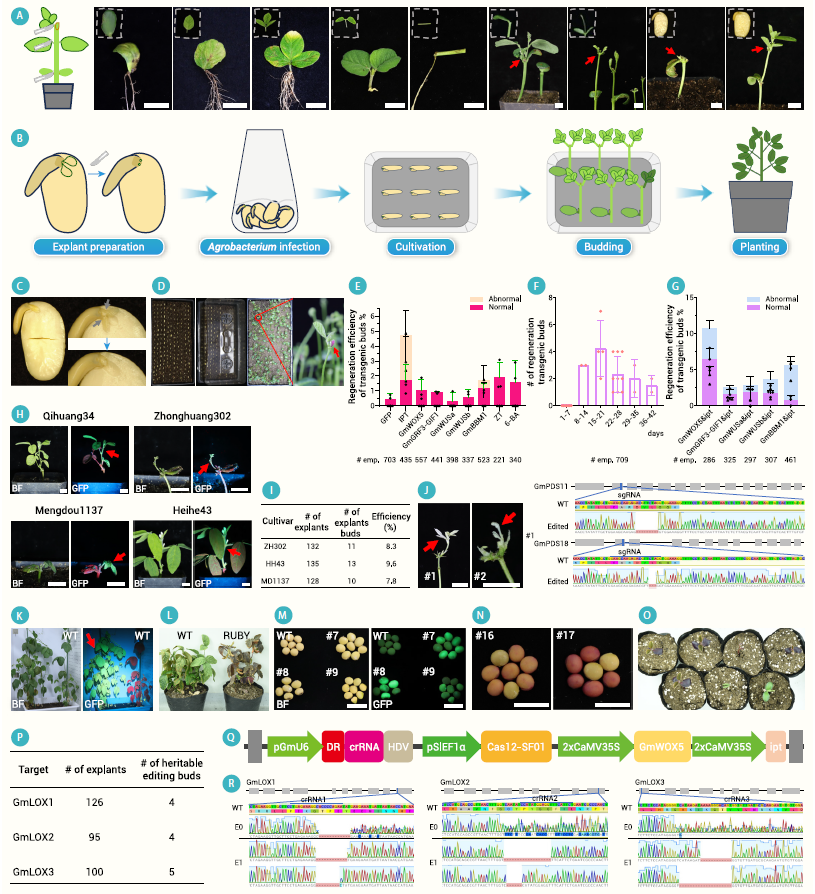
图9 大豆CDB遗传转化方法的建立(Cao et al., 2025)。(A)在无组织培养条件下,大豆离体和原位组织的器官再生。从左至右依次为:外植体取样示意图,子叶叶柄、真叶叶柄、复叶叶柄、茎、下胚轴、子叶腋区(6日龄幼苗)、初生叶(6日龄幼苗)、子叶腋区(吸胀种子)、胚芽(吸胀种子),红色箭头指向再生芽;(B)大豆CDB转化流程示意图;(C)外植体照片,灰色箭头指向切割位点;(D)外植体移栽至土壤中生长;(E)不同发育调控因子在无组织培养条件下对大豆再生效率的影响;(F)大豆CDB系统所需的再生时间;(G)不同发育调控因子组合在无组织培养条件下对大豆再生效率的影响;(H)四个大豆品种中表达GFP的幼苗照片;(I)利用CDB结合GmWOX5 和ipt 对三个大豆品种进行转化的效率;(J)齐黄34中PDS 基因编辑后白化芽的代表性照片和Sanger测序结果;(K)表达GFP的齐黄34植株与对照(WT)的生长情况;(L)表达RUBY的齐黄34植株与对照(WT)的生长情况;(M)表达GFP的植株所结的种子;(N)表达RUBY的植株所结种子;(O)表达RUBY的种子长成的幼苗;(P)利用CRISPR-Cas12-SF01对GmLOX1 /2 /3 基因进行可遗传编辑的统计数据;(Q)Cas12-SF01基因编辑载体示意图;(R)经Cas12-SF01编辑GmLOX1 /2 /3基因的E0和E1植株的Sanger测序结果。
文章至此就告一段落了!在本次推文中小远接着前期的内容,又给大家介绍了四种快速、高效的遗传转化方法,希望对大家的转化工作有一定的启发。最后,再插播一下广告,我司目前可以为大家提供大豆、番茄、烟草等四十大物种的遗传转化服务,有需要的小伙伴可以查询我司官网或致电详询哦!
References:
Cao X, Xie H, Wang Z, et al. An efficient tissue-culture-free soybean genetic transformation technology using the extremely simple cut-dip-budding strategy. Innovation (Camb). 2025, 7(3): 101221.
D'Amico F, Candela M, Turroni S, et al. The Rootstock Regulates Microbiome Diversity in Root and Rhizosphere Compartments of Vitis vinifera Cultivar Lambrusco. Front Microbiol. 2018, 9: 2240.
Iwase A, Harashima H, Ikeuchi M, et al. WIND1 Promotes Shoot Regeneration through Transcriptional Activation of ENHANCER OF SHOOT REGENERATION1 in Arabidopsis. Plant Cell. 2017, 29(1): 54-69.
Kshetry AO, Ghose K, Alok A, et al. A synthetic transcription cascade enables direct in planta shoot regeneration for transgenesis and gene editing in multiple plants. Mol Plant. 2025, 18(12): 2066-2081.
Li Y, Lu K, Feng S, et al. ROTIS: A RUBY-assisted one-shot tissue-culture-free system for in-planta soybean transformation from germinated seedlings. Plant Physiol. 2025, 199(4): kiaf609.
Song J, Sun P, Kong W, et al. SnRK2.4-mediated phosphorylation of ABF2 regulates ARGININE DECARBOXYLASE expression and putrescine accumulation under drought stress. New Phytol. 2023, 238(1): 216-236.
Zhang Y, Ming R, Khan M, et al. ERF9 of Poncirus trifoliata (L.) Raf. undergoes feedback regulation by ethylene and modulates cold tolerance via regulating a glutathione S-transferase U17 gene. Plant Biotechnol J. 2022, 20(1): 183-200.
Zhang SY, Luo RF, Wu YX, et al. Establishment and application of high-pressure propagation breeding (HPPB)-mediated genetic transformation system in citrus rootstocks. Plant Biotechnol J. 2025, 23(7): 2790-2792.