本文内容速览:

小远在"育种家看过来------利用基因编辑快速创制雄性不育系"一文中给大家介绍过关于雄性不育 及杂种优势 的概念,以及利用基因编辑技术创制不育系的方法,大家在阅读本文之前可以先回顾一下。利用杂种优势生产的杂交种,目前在我国农业生产中占据举足轻重的地位------尤其是在玉米中,杂交种覆盖率已接近100%。然而,要高效利用杂种优势,就必须解决母本去雄这一核心环节。此前我们讲到的基因编辑技术 擅长解决的是 "如何创制不育系本身"的源头问题 ,而今天要介绍的转基因技术 ,则聚焦于"如何繁殖和筛选不育系"的配套问题。接下来我们以玉米为例进行说明。
在玉米杂交种生产中,母本去雄是最耗费人力与成本的关键环节。人工去雄窗口期短、劳动密集;机械去雄虽已有初步应用,但现有设备去雄率约90%,仍需人工二次去雄,且易损伤叶片造成减产;化学杀雄效果不稳定,难以满足商业化纯度要求。一个理想的解决方案是让母本在遗传上实现雄性不育。
P art.01
雄性不育的分类
玉米雄性不育根据遗传机制可分为两类:细胞质雄性不育(Cytoplasmic Male Sterility,CMS ) 与细胞核雄性不育(Genic Male Sterility,GMS)。
细胞质雄性不育(CMS)
CMS是受细胞质遗传基因(即线粒体不育基因)与细胞核育性恢复基因共同决定。根据败育发生的分子机制和对育性恢复基因的响应差异,玉米CMS可进一步分为CMS-T、CMS-C和CMS-S三种主要类型,三种类型分别对应不同的不育胞质类型及不同的恢复基因调控方式(Beckett, 1971)。
CMS-T是玉米CMS研究中最早被克隆和解析的类型。T型CMS的不育性由线粒体嵌合基因T-urf13引起,该基因编码的urf13蛋白能够形成孔道结构导致花粉败育,同时该蛋白也是玉米小斑病T小种毒素的受体,解释了CMS-T不育系易感小斑病的深层原因。
CMS-C是我国玉米育种中应用较广的不育类型,其不育由线粒体嵌合基因atp6-c等异常转录本介导,同时多个核恢复基因参与其育性恢复调控。
CMS-S的不育机制更为复杂,在株系水平上育性表现不稳定,常受环境因素影响而出现部分可育或育性波动现象。育种实践表明,CMS-S的育性波动是限制其在大规模制种中应用的重要原因之一。
CMS已通过"三系配套"实现了一定的规模化应用,所以它们应用的核心难题并非无法自我繁殖,而是胞质单一 带来的遗传脆弱性风险 、恢复系 来源有限 、 以及部分类型育性不稳定等限制了其应用潜力。
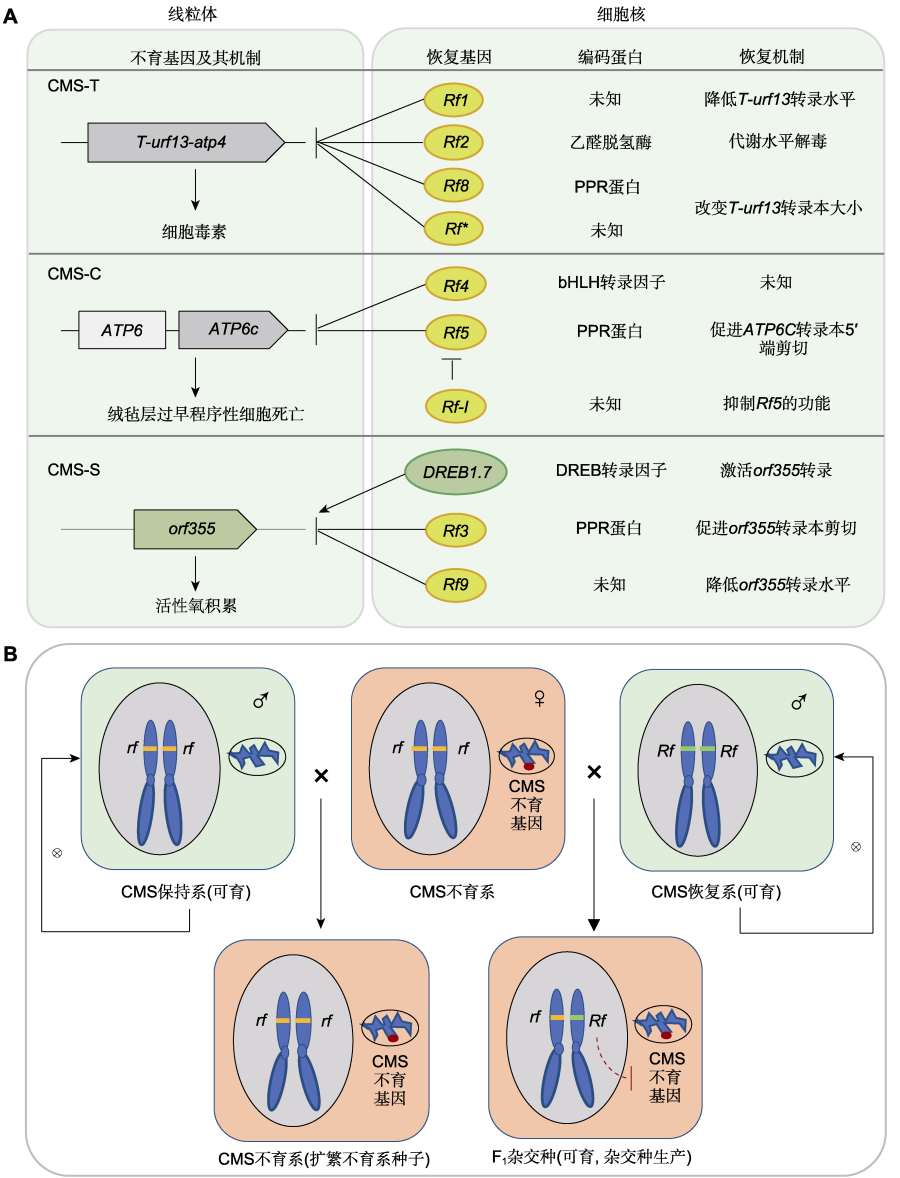
图1 玉米细胞质不育(CMS)和三系法杂交制种(吴锁伟等, 2024)。(A)三种类型细胞质不育及其育性恢复机制(箭头表示激活作用,T型线表示抑制作用); (B)三系法杂交制种原理。
细胞核雄性不育(GMS)
GMS的不育性状受细胞核基因控制, 且表现出典型的孟德尔遗传。按照不育性状的显隐性关系,可进一步分为隐性核不育和显性核不育两种类型。目前发现的玉米核不育材料大部分为隐性核不育,仅少数几个为显性核不育(吴锁伟和万向元, 2018)。根据育性是否受环境影响, 细胞核雄性不育还可分为两类,一类为育性不受环境影响的GMS, 另一类为育性受光照时长和温度等环境影响的GMS, 即光温敏核雄性不育(P/TGMS),一般表现为长日照不育、短日照可育以及高温不育、低温可育(吴锁伟等, 2024)。
GMS的致命缺陷在于不育系无法自我繁殖:当不育株(msms)与杂合可育株(Msms)杂交时,后代以1:1的比例分离出不育株与可育株,且二者从种子上无法区分。这一难题长期阻碍了GMS的商业化应用。不过,技术手段可以弥补这一不足------为了解决纯合不育系的持续繁殖和种子分选问题,转基因技术被引入GMS领域。
P art.02
前期理论基础
1993年:转基因保持系的概念验证
转基因技术最早应用于核不育保持系的构建可以追溯到1993年。Plant Genetic System公司的Williams Mark 和 Leemans Jan在这一年率先尝试了利用基因工程解决核不育问题的初步方案,并发表了一篇题为"Maintenance of Male-sterile plants"的专利,可惜因为年代久远,小远没能下载到原文。如果大家有办法找到这篇专利,非常欢迎跟小远分享一下!
该公司的核心思路是:在纯合雄性不育植株(msms )中转入育性恢复 基因与花粉败育 基因。通过这样的遗传设计,获得的保持系可以通过自交同时产生不育系种子与保持系种子------不育系种子来自不携带转基因构件的配子组合(msms );保持系种子则来自携带转基因配子的组合(Msms)。因此,在操作层面上,核不育系实现了可持续的自我维持。
然而,这一早期方案存在一个致命的实际操作瓶颈:不育系种子与保持系种子的外观完全相同,无法通过物理手段高效分选。换句话说,育种者在繁殖世代不得不将混合种子全部播种,然后根据田间植株的育性表现(即花期辨别可育株与不育株)进行人工鉴别和淘汰。这一过程虽然解决了核不育系从"不可繁殖"到"可繁殖"的基本困境,但田间鉴别的人工成本仍然居高不下,极大限制了该技术方案的规模化应用。
尽管如此,1993年的工作仍具有重要的开创性意义:它第一次证明了通过转基因手段创制核不育系保持系在技术上是可行的,为后续所有商业化转基因核不育制种技术提供了最基础的操作范式和验证数据。
2002年:Perez-Prat的理论飞跃
如果说1993年的实践为核不育转基因制种提供了"怎么做"的技术验证,那么2002年来自Aventis CropScience N.V.公司的Perez-Prat和Van Lookeren Campagne在Trends in Plant Science杂志上发表的一篇题为"Hybrid seed production and the challenge of propagating male-sterile plants"的文章则为这个方向提供了"怎么做才更好"的理论重构。
两位科学家敏锐地指出:前面提到的早期技术方案之所以难以落地,根本症结不在于转基因设计本身,而在于不育/可育种子的筛选效率过低。单纯依靠转基因保持系实现核不育系的自我繁殖是不够的------育种者还需要一种高效的方法,在不播种的情况下就能区分两类种子。
他们的理论解决方案是:在转基因构件中增加第三个功能元件------报告基因(图2)。该报告基因在种子阶段特异性表达,赋予转基因保持系种子某种肉眼可辨或被机器识别的表型特征(例如胚乳颜色或荧光信号)。这样一来,不育系种子(不含转基因)与保持系种子(含转基因)之间就可以通过机械化颜色分选高效分离,彻底告别田间人工鉴别的低效时代。
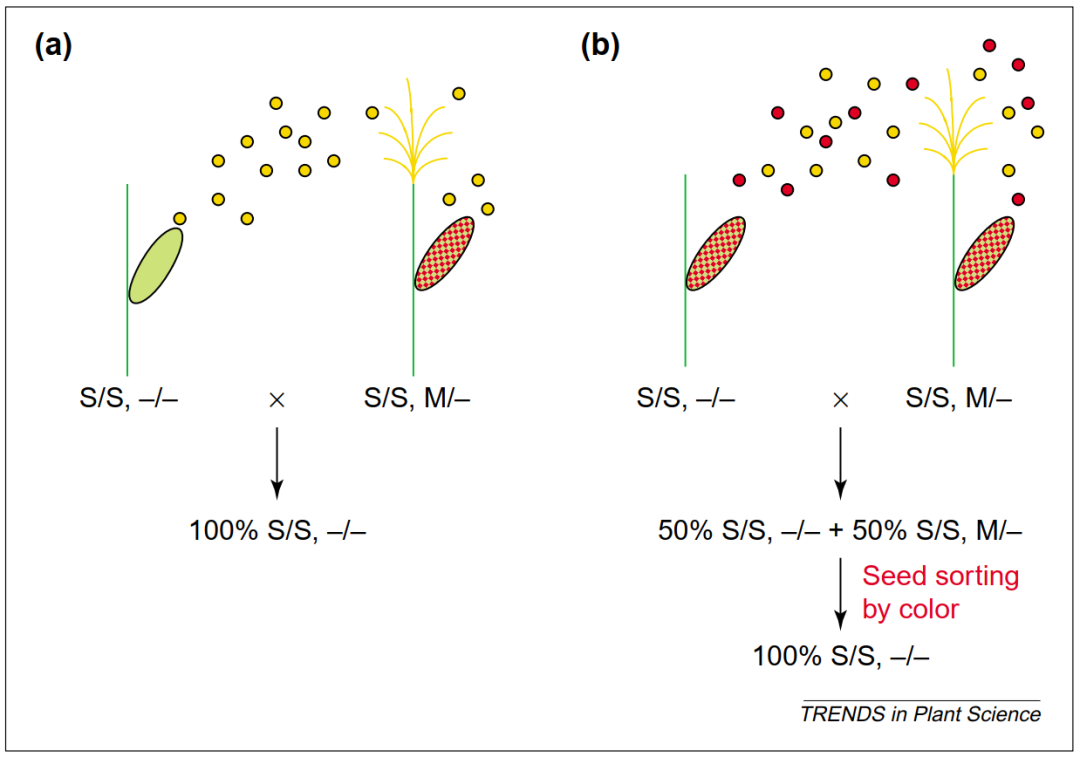
图2 保持系繁殖雄性不育系(Perez-Prat and Van Lookeren Campagne, 2002)。 以两种不同的玉米保持系为例:(a) 花粉致死型(barnase )保持系和 (b) 颜色标记型(C1-S 和 B-peru )保持系。两种情况下,保持系与雄性不育系的遗传背景完全相同,唯一的区别是保持系含有一个转基因维持位点 ( M **)**,该位点使其雄性可育(a)花粉致死型保持系植株只产生 S,- 花粉粒(黄色),因为携带 S,M 的花粉粒被 M 位点中的花粉致死基因杀死。因此,雄性不育系(S/S, -/-)与花粉致死型保持系(S/S, M/-)杂交的后代全部为雄性不育(S/S, -/-);(b)颜色标记型保持系可同时产生有活力的 S,-(黄色)和 S,M(红色)花粉粒。雄性不育系(S/S, -/-)与颜色标记型保持系(S/S, M/-)杂交的后代中,M 位点按 1:1 分离,但携带该位点的种子带有颜色。通过分拣种子并淘汰有色种子,即可获得全部为雄性不育的后代。
这一"报告基因辅助分选"理论的提出,被广泛认为是转基因核不育制种技术史上的分水岭。它让育种家意识到:核不育制种体系的"保持系"不只是一个基因型的载体,更是一个"自带标签"的智能化系统,能够通过简单的物理筛选实现不育系与保持系的高效分离。此后,所有商业化的转基因核不育制种技术------无论是SPT、RHS2,还是SSMM/SMART------其设计理念都直接或间接地受到了2002年这篇观点文章的理论影响。接下来,我们将具体探讨这些商业化的转基因核不育制种技术,并同时介绍一种尚未商业化的相关技术------MCS。
P art.03
转基因核不育制种技术
玉米SPT技术
种子生产技术(Seed Production Technology,SPT)是首个真正实现商业化应用的转基因核不育制种系统,由美国杜邦先锋公司在2006年于玉米上实验成功。2012年,SPT技术生产的杂交种子在美国上市,并在美国、日本、澳大利亚、新西兰等国家取得非转基因农产品认证,实现商业化。
2016年,杜邦先锋公司的科研团队在Plant Biotechnology Journal杂志上发表了一篇题为"Development of a novel recessive genetic male sterility system for hybrid seed production in maize and other cross-pollinating crops"的文章,系统阐述了SPT技术的完整分子设计(图3)和商业化验证数据(图4)。
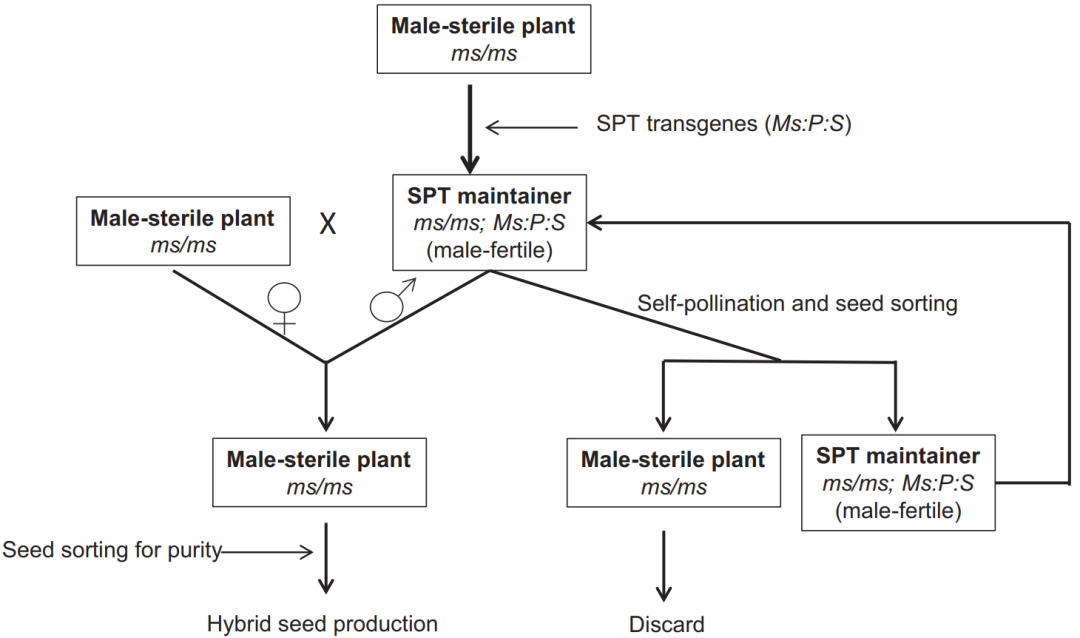
图3 用于扩繁隐性核雄性不育(ms )系的系统示意图(Wu et al., 2016)。Ms :育性恢复基因;P :花粉致死基因;S:种子可筛选标记基因;SPT:种子生产技术。
SPT保持系的转基因构建体包含三个核心功能元件:①Ms45 育性恢复基因:携带Ms45 野生型等位基因,互补玉米隐性核不育突变ms45 的功能缺失,使保持系植株恢复可育状态;②α-淀粉酶基因(zm-aa1 ): 在花粉中特异性表达,降解淀粉粒,使携带转基因的花粉丧失萌发能力------这是"花粉败育"的功能模块,防止转基因通过花粉向环境扩散;③DsRed2红色荧光蛋白报告基因:在种子中特异性表达,赋予转基因保持系种子红色荧光表型。
SPT保持系植株由于携带了Ms45 育性恢复基因,自身能够正常产生花粉并完成自交结实。但在自交过程中,花粉的分离呈现一个不对称特征------携带转基因构件(含α-淀粉酶和DsRed2 )的花粉因α-淀粉酶降解淀粉而丧失萌发能力,只有不携带转基因的花粉才能参与授精。因此,SPT保持系的自交后代产生两种种子:一半是非转基因的不育系种子(msms ,无转基因,无DsRed2红色荧光),另一半是转基因的保持系种子(Msms,含完整的转基因元件,种子呈红色荧光)。由于两类种子可通过红色荧光信号被机械化颜色分选仪高效分离,育种者可以轻松获得100%的不育系种子用于杂交制种,同时保留部分保持系种子用于下一轮繁殖(图4)。这种方法创制出来的不育系当作母本用于杂交制种后,子一代杂交种不携带转基因成分。
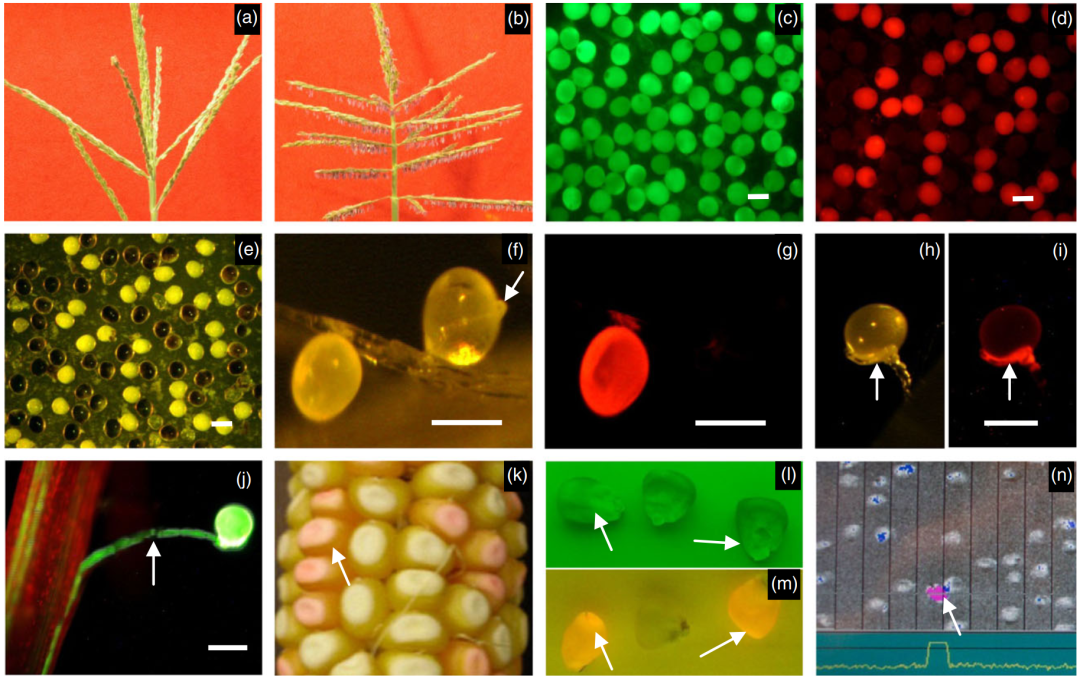
图4 转基因植物的特征分析(Wu et al., 2016)。(a)ms45 植株的不育雄穗;(b)含有 pPG47::Bt1:zm-aa1// pMs5126::Ms45// p35S::PAT 的转基因 ms45 植株的可育雄穗;(c-i)含有 pPG47::Bt1:zm-aa1//35 S EN- pUbi::PAT::DsRed-Express 的转基因植株的花粉。(c、d)用荧光素二乙酸酯染色的成熟花粉(c),以及在红色荧光蛋白滤镜下观察的同一花粉(d)。(e) 用碘化钾染色的成熟花粉样本(与 c 相同样本)。(f-i)体内花粉萌发实验。(f、g) 无花粉管的 DsRed-Express 花粉 与 有花粉管的非 DsRed-Express 花粉,分别在可见光(f)和 DsRed-Express 滤镜下(g)观察。(h、i)一个 DsRed-Express 花粉粒的异常花粉管生长,分别在可见光(h)和 DsRed-Express 滤镜下(i)观察;(j) 含有 pUbi::YFP 的对照转基因植株的花粉管生长。图 f、h‑j 中的箭头表示花粉管。(k-n)种子中 DsRed2 表达及种子分选特征。(k)含有 pMs5126::Ms45// pPG47::Bt1:zm-aa1//35SEN- p*LTP2::DsRed2(Alt1)*的转基因植株的果穗。(l、m)非 DsRed2(黄色)和 DsRed2(红色)种子分别在绿色滤镜(l)和红色滤镜(m)下观察。(n) 在种子分选机上流动的种子(从上到下)。图 k‑n 中箭头指向 DsRed2 种子。
玉米RH2技术
与SPT依赖于"种子颜色分选"的方案不同,孟山都采取了另一条技术路线------草甘膦诱导型雄性不育系统。利用多数作物的雄配子对草甘膦的敏感性, 在合适的时期喷施草甘膦会导致植物雄性不育,第一代RHS(RHS1)基于CP4 EPSPS 基因在花粉中的低表达水平:CP4 EPSPS是对草甘膦不敏感的5-烯醇丙酮酸莽草酸-3-磷酸合酶,当施草甘膦时,雄蕊组织因CP4 EPSPS表达水平不足而被草甘膦灭杀,从而诱导雄性不育。然而,RHS1系统的育性诱导效率在部分自交系中存在不稳定问题。
2013年,孟山都公司成功开发了第二代草甘膦诱导型雄性不育系统(RHS2),其核心技术创新在于将RNA干扰(RNAi)机制与CP4 EPSPS 基因相结合。2018年,孟山都科研团队在PLOS ONE杂志上发表了一篇题为"Endogenous tassel-specific small RNAs-mediated RNA interference enables a novel glyphosate-inducible male sterility system for commercial production of hybrid seed in Zea mays L."的研究论文,系统阐述了RHS2技术的分子设计原理及其作用机制(图5)。
RHS2在转基因构建体中引入了玉米内源雄穗特异性小干扰RNA(mts-siRNAs)的靶序列,使CP4 EPSPS的mRNA在雄穗组织中被特异性切割,从而在雄穗中实现CP4 EPSPS蛋白的靶向"沉默"------雄穗缺乏CP4 EPSPS保护,在草甘膦处理下被选择性灭杀;而营养组织和雌蕊因CP4 EPSPS正常表达而耐受草甘膦。
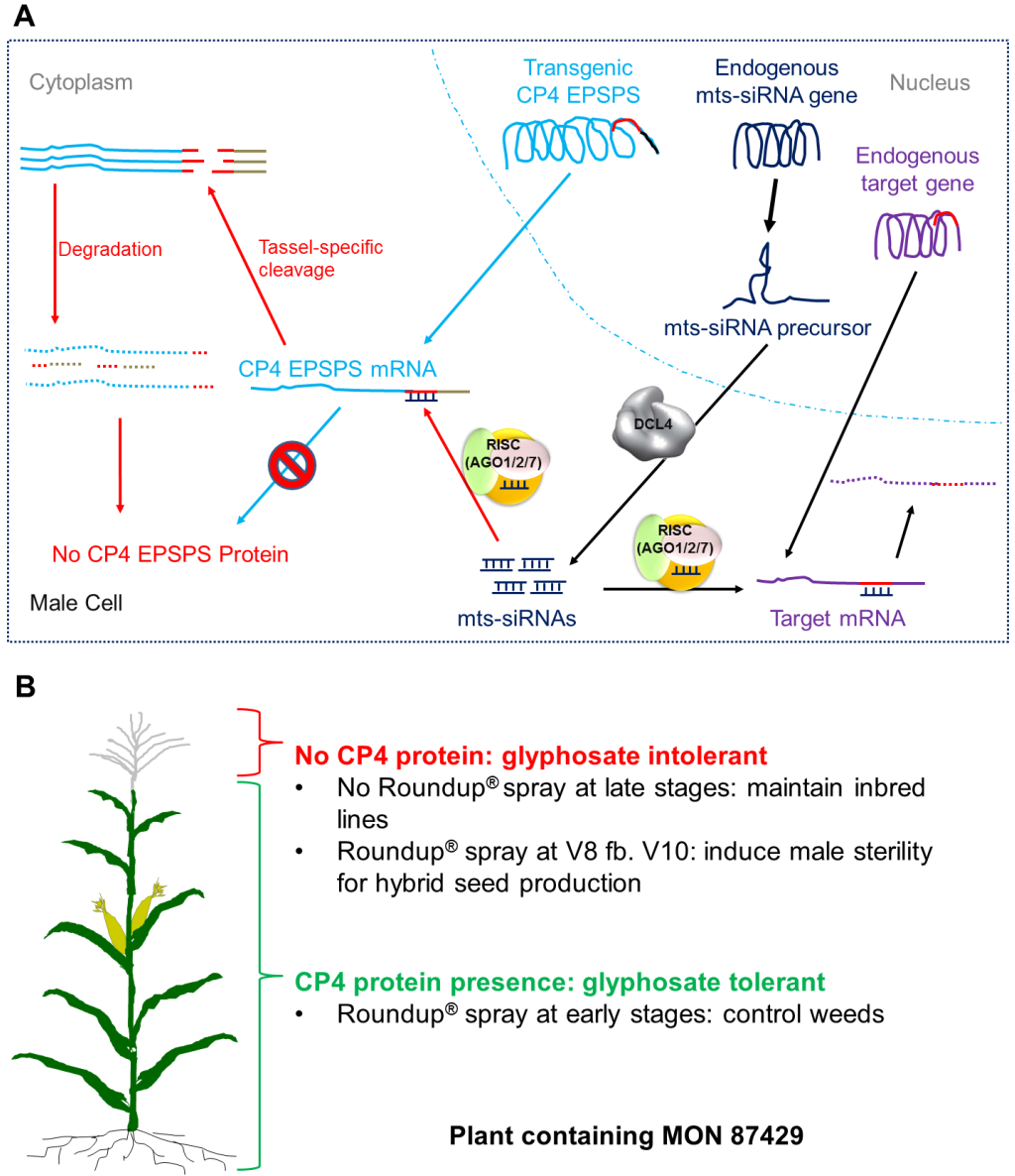
图5 RHS2技术(Yang et al., 2018)。(A)分子作用模式。内源的mtssiRNA(雄穗特异性小干扰RNA)触发CP4 EPSPS mRNA的切割,这一过程很可能由AGO2蛋白介导,且仅发生在雄性生殖细胞中,具有序列特异性。切割产物由于失去了3′端的poly(A)尾和5′端的帽子结构的保护,分别被3′→5′和5′→3′方向的核糖核酸外切酶降解。结果导致雄性生殖细胞中几乎没有CP4 EPSPS蛋白合成;(B)双重用途。CP4 EPSPS蛋白在营养组织和雌性组织中正常积累,为早期季节喷施草甘膦进行杂草控制提供了出色的保护,因此该自交系可以被繁殖。相比之下,雄性生殖细胞中几乎没有CP4 EPSPS蛋白。当在花粉发育的关键阶段喷施草甘膦时,雄性不育被诱导,这些植株可作为母本用于生产杂交种子。缩略词:AGO---Argonaute蛋白;DCL---Dicer样蛋白;RISC---RNA诱导的沉默复合体。未喷草甘膦或仅早期喷:植株整体绿色(耐草甘膦),雄穗正常散粉,雌穗正常结籽→用于自交系繁殖;在V8---V10喷施草甘膦:营养组织仍绿色(耐),但雄穗枯萎不育(红色叉号),雌穗正常→用于杂交制种。
RHS2的最大优势在于无需保持系繁殖和种子颜色分选。在杂交制种过程中,育种者将转基因母本植株与常规父本自交系混种,并在玉米进入雄穗发育关键阶段后,整田喷施草甘膦。母本的雄穗因缺乏CP4 EPSPS保护而被草甘膦灭杀,实现雄性不育;而营养组织和雌蕊因CP4 EPSPS正常表达而保持健康。母本接受父本花粉后即可生产杂交种。RHS2中使用的mts-siRNAs在不同玉米种质间高度保守,这使得该系统的诱导不育效果可在广泛的自交系背景中复制。但由于母本是转基因植株,其生产的杂交种一般也会携带转基因成分。
玉米SSMM技术和玉米MCS技术
玉米智能核不育制种技术(Smart Seedling Male Sterility,SSMM)由中国农业大学赖锦盛教授团队历经十余年研发,2025年入选农业农村部"农业重大引领性技术"。与SPT类似,SSMM也采用了"颜色/形态标记+机械化分选"的策略,但在标记的丰富性和系统集成性上有所突破,当保持系与核不育系杂交后,不育系种子与保持系种子之间会表现出显著的籽粒大小差异和植株颜色差异,使得分选不再局限于荧光检测环节。目前尚未检索到该技术细节的研究论文,不过研究团队已获得5项中国和2项美国专利授权,有兴趣的朋友可自行查阅了解。
玉米多控不育技术(Multi-control Sterility System,MCS)由北京科技大学生物前沿技术与应用研究中心万向元团队研发。该技术将玉米花粉育性恢复基因(ZmMs7 )、花粉自我降解基因(ZmAA 、 Dam )、红色荧光蛋白基因(Ds R ed2 或mCherry )和抗除草剂基因(bar)组合在一起,构建多控不育遗传转化载体,通过遗传转化和杂交、回交转育技术,导入到玉米隐性核雄性不育系中,以恢复不育系的育性并能有效繁殖,实现一系两用的目的(图6)。
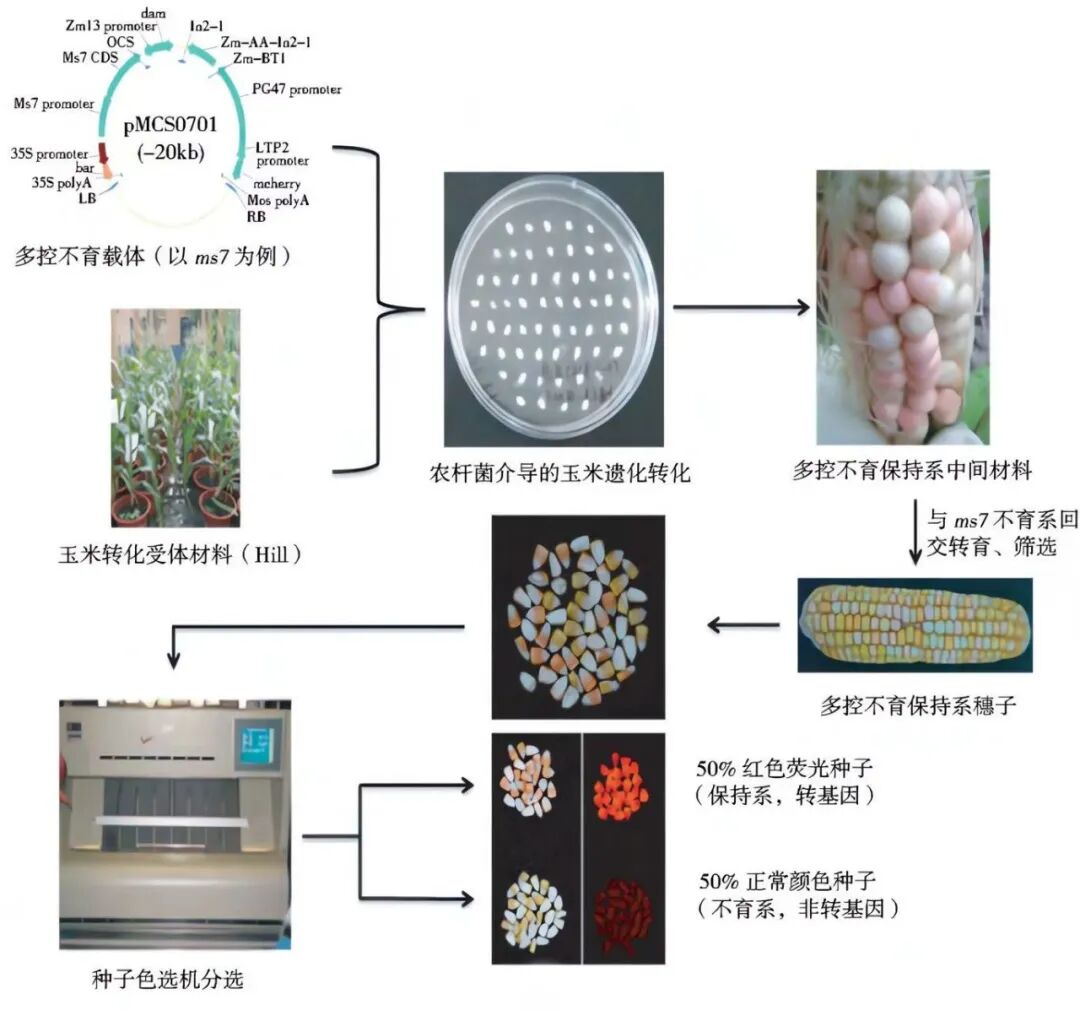
图6 玉米多控不育技术体系的整体技术路线(吴锁伟和万向元, 2018)
在转基因核不育制种的技术框架下,没有唯一的"最优解",不同方案适应不同的产业需求与监管环境。小远将本篇内容给大家总结成了一个表格,便于大家直观的比较,上述这些技术路线虽然各具优势,但都依赖转基因手段构建保持系,这意味着它们需要经过复杂的转基因安全评估和监管审批程序,在某些国家和地区可能面临推广壁垒。近年来基因编辑技术在飞速的发展,利用基因编辑技术与转基因报告基因系统结合的"一步法制种"策略为创制新的核不育体系提供了可能。如果大家感兴趣的话,可以给小远留言,我们去挖掘一下目前"一步法制种"策略在各个物种中的进展。
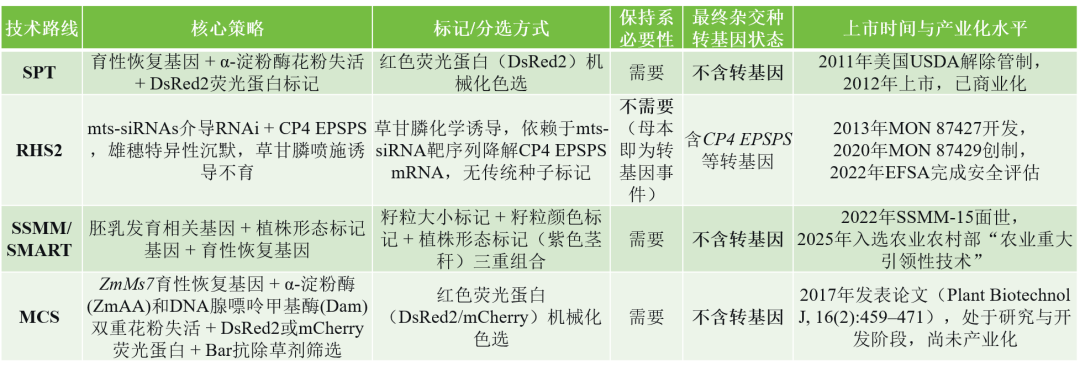
表1 主要GMS商业化技术路线的核心策略、标记方式、转基因状态及产业化进展
References:
吴锁伟, 安学丽, 万向元 (2024). 玉米雄性不育机理及其在工程核不育制种中的应用. 植物学报 59, 932--949.
吴锁伟****,**** 万向元 (2018). 利用生物技术创建主要作物雄性不育杂交育种和制种的技术体系. 中国生物工程杂志 38, 78--87.
Beckett JB (1971). Classification of male-sterile cytoplasms in maize (Zea mays L.). Crop Sci11, 724--727.
PLANT GENETIC SYSTEMS N.V.."MAINTENANCE OF MALE-STERILE PLANTS."CA2137559.2005-11-22.
Perez-Prat E, van Lookeren Campagne M M. Hybrid seed production and the challenge of propagating male-sterile plantsJ. Trends in Plant Science, 2002, 7(5): 199-203.
Wu Y, Fox T W, Trimnell M R, et al. Development of a novel recessive genetic male sterility system for hybrid seed production in maize and other cross‐pollinating cropsJ. Plant Biotechnology Journal, 2016, 14(3): 1046-1054.
Yang H, Qi Y, Goley M E, et al. Endogenous tassel-specific small RNAs-mediated RNA interference enables a novel glyphosate-inducible male sterility system for commercial production of hybrid seed in Zea mays LJ. Plos one, 2018, 13(8): e0202921.