阐明转录因子(TF)的功能,对于深入理解和调控协调植物生长、发育及抗逆性的基因表达程序机制至关重要。确定转录因子的基因组结合位点------即顺式调控组(cistrome)------尤为关键,因为它揭示了转录因子发挥作用的具体位置,并且当与基因表达(RNA-seq)和染色质景观数据(ATAC-seq)相结合时,能够全面勾勒出转录因子的功能范。
本期,我们介绍一篇近期发表在Genome Biology上的综述《Many roads lead to a plant cistrome: mapping and interpreting transcription factor binding in plants》。这篇综述主要讨论了如何在植物中绘制和解析转录因子结合位点图谱,谈了目前主流的技术以及影响天然cistrome的因素。

1 什么是顺式调控组(cistrome)?
这篇综述把cistrome定义为**某一个转录因子在特定基因组中的全部DNA结合位点集合。**也就是说,如果一个TF在基因组上能结合到若干启动子、增强子或其他顺式调控区域,那么这些结合位点加起来就是它的cistrome。
文章提出有多个层面的的cistrome。
0 1 Native cistrome(天然cistrome)
Native cistrome指在真实体内、某一细胞类型、某一环境和某一时间点下,一个TF实际发生的全部DNA结合事件。这是最接近生物真实状态的cistrome。但文章强调native cistrome实际上无法被完全无扰动地测到。因为从取样、交联、细胞核提取、抗体捕获、建库、测序到peak calling,每一步都会改变或筛选原始结合事件。
0 2 Observable cistrome(可观测cistrome)
Observable cistrome这是实验技术最终检测到的那部分TF结合位点。它不是native cistrome的完整复制,而是经过实验和分析流程筛选后的结果。影响因素包括交联效率、细胞核/染色质提取效率、抗体特异性、染色质打断效率、测序深度、peak caller选择、FDR阈值、归一化方法等。
0 3 Observable functional cistrome(可观测功能cistrome)
不是所有检测到的TF结合位点都有功能。functional cistrome指的是在observable cistrome中,真正影响基因表达、参与调控输出的那一部分结合位点。例如综述提到,早期拟南芥转录因子EIN3 ChIP-seq研究中,只有约三分之一EIN3结合位点伴随转录响应。所以 " 有peak " 不等于 " 有功能 " 。
2 植物cistrome mapping的主要技术路线
综述中重点介绍了四种主流的技术ChIP-seq、CUT&RUN、CUT&Tag和DAP-seq技术。
ChIP-seq
经过过去二十年的持续改进,ChIP-seq仍是基因组顺式作用元件图谱构建的金标准**。**优点有技术成熟,文献积累多,可用于TF、组蛋白修饰、RNA Pol II、染色质因子等,仍是很多研究的基准方法。缺点是对抗体依赖强、输入材料通常较多、植物组织提取复杂、分辨率相对有限,峰宽常在200-400bp,交联和过表达标签TF可能带来偏差。文章还列举了很多基于ChIP的植物优化改进技术,比如eChIP、PHILO ChIP-seq、TARGET/μChIP-seq、aChIP等。

图1:植物中四种主要顺式作用元件图谱构建方法的示意图概述
CUT&RUN
CUT&RUN是低背景、高分辨率的替代方案,它的思路是用抗体定位转录因子,再用pA/G-MNase在结合位点附近切割DNA,释放目标片段后测序。优点背景低、输入量少、分辨率高于传统ChIP-seq、不需要大规模染色质超声打断。
CUT&Tag
CUT&Tag与CUT&RUN类似,但使用的是pA/G-Tn5。Tn5会在目标位点附近同时完成切割和接头插入,因此流程更快。优点有输入材料少、灵敏度高、背景低、建库流程简化。潜在问题是Tn5偏好开放染色质,因此开放区域可能在数据中被过度富集。此外,针对植物CUT&Tag也有一些拓展方法,比如nCUT&Tag、snCUT&Tag、B-CUT&Tag、tsCUT&Tag。
DamID-seq和ChEC-seq
综述也提到很多TF-DNA结合是瞬时的,传统ChIP/CUT类方法可能漏掉。文章提到了两个技术:DamID-seq和ChEC-seq,它们可以捕捉到瞬时的结合。DamID-seq把TF与大肠杆菌Dam甲基转移酶融合,后者可对TF-DNA互相作用位点附近的GATC基序的腺嘌呤残基进行甲基化,从而在TF靶启动子处留下稳定且可遗传的标记。优点是可以捕捉短暂结合;缺点包括依赖GATC分布、TF-Dam融合可能改变TF行为、可能产生异位结合。ChEC-seq(酵母应用较为广泛)是将TF与MNase融合,通过诱导切割定位结合位点。不过,该技术植物中应用还较少。
DAP-seq
DAP-seq是植物领域非常重要的体外cistrome方法,也是综述重点强调的一大技术。流程是体外表达重组TF,与打断的基因组DNA孵育,亲和纯化TF-DNA复合物,测序得到结合位点。优点是不需要转基因植物,不依赖体内抗体,可大规模、高通量应用,适合非模式植物和作物 ,已用于拟南芥、玉米、番茄、银杏等多种植物。缺点是DAP-seq测的是体外cistrome不是体内真实结合图谱 。它缺少真实TF剂量信息,染色质结构,核小体阻挡,组蛋白修饰背景,辅因子,细胞类型和环境信息。因此DAP-seq往往得到比体内方法更多的peaks,因为很多体内被染色质遮挡的区域在体外可以被TF结合。不过基因组DNA保留了其天然DNA甲基化信息,因此DAP-seq可用于研究TF对DNA甲基化的敏感性。文章提到ampDAP-seq使用无甲基化DNA,可揭示甲基化对TF结合的抑制作用。
尽管存在这些局限性,DAP-seq技术的简便性、可扩展性及广泛适用性仍远胜于其体外实验的局限 ------尤其是在将DAP-seq数据与开放染色质及基因表达数据(ATAC-seq和RNA-seq)相结合时,可更精准地定位具有功能相关性的结合位点。
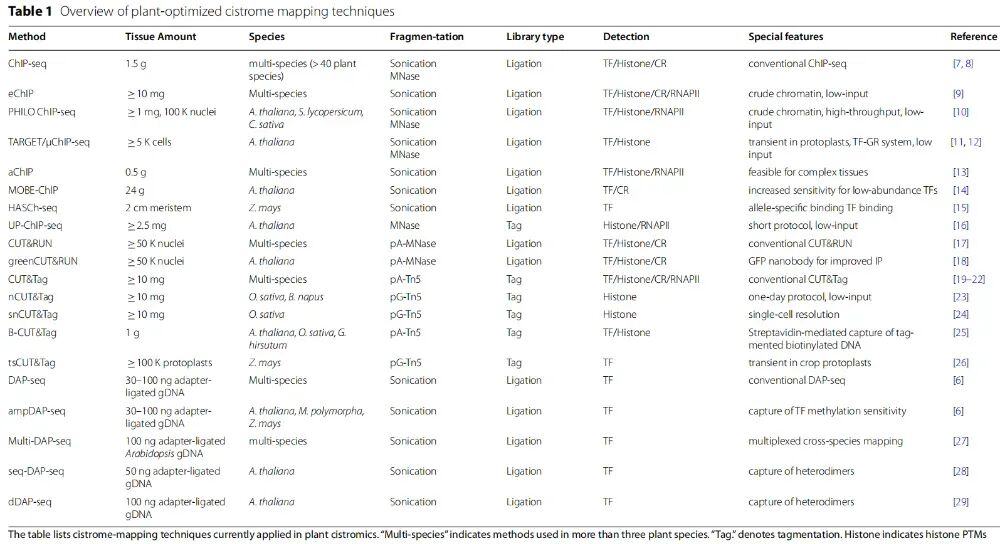
针对植物优化的cistrome图谱绘制技术概览
Total cistrome mapping
Total cistrome mapping是指看所有TF的总体图谱。但单个TF一个个很难测,因为植物中通常有约2000个TF,而且抗体稀缺。综述中介绍了一些间接测总cistrome的方法 ,比如MOA-seq、ATAC-seq footprinting(可识别开放染色质区域,TF结合位置由于被保护,会出现局部信号缺口,可用于推断TF的footprint)、SAM-seq和seq-ATI。
3 天然cistrome形成因素
天然cistrome包含特定细胞和环境背景下的所有体内转录因子(TF)结合事件。原封不动地捕获天然顺反子组是不可能的,因为其组成在实验流程的每个阶段都会发生重塑------从TF-DNA结合事件,到抗体捕获的TF-染色质复合物,最终到基因组浏览器中观察到的峰。文章认为,native cistrome的形成主要由四层因素决定:TF剂量、染色质环境、辅因子互作和顺式调控元件结构。
0 1 转录因子剂量(TF dosage)
TF dosage指的是某个时间点细胞核中能够真正结合DNA的有效TF数量。低剂量时,TF优先结合高亲和力位点,开放染色质位点。剂量升高后,TF可能扩展到低亲和力位点,较不开放区域以及非典型结合位点。因此,TF过表达会显著改变cistrome。文章列举了很多例子,番茄Woolly转录因子的剂量控制毛状体分化;拟南芥的转录因子PLT1/PLT2以剂量依赖方式控制根发育;WUS剂量梯度维持茎尖分生组织干细胞状态;MYC2的cistrome会随环境动态变化。
0 2 辅 因子互作 ( Cofactor interactions )
TF是否能结合DNA,不仅取决于自己,还取决于辅因子。比如能够形成异源二聚体的转录因子,以及能够隔离或抑制转录因子的蛋白质。比如,MYC2可与MTB1/2/3形成异二聚体,从而阻碍MYC2 DNA结合。
0 3 染色质环境(Chromatin environment)
染色质状态会决定某些motif是否可被TF接近。影响因素包括核小体占据,染色质开放程度,H3K4me3、H3K27ac等活跃组蛋白标记,H3K27me3、H3K9me2/3 等抑制标记,以及DNA甲基化。
0 4 顺式调控元件结 构(CRE architecture)
由DNA序列影响的CRE结构也会影响cistrome,即使是很小的遗传变异,比如SNP或indel,也可能改变CRE的组成、间距、方向或表观遗传状态,从而重塑TF的结合位点分布,也就是改变cistrome。比如,自然群体或不同物种之间的CRE序列差异,会导致TF结合位点发生变化。文章中举了案例,玉米两个自交系的DAP-seq比较对30个TF家族的200个cistrome进行比较,发现两种玉米自交系之间存在大量结合差异,主要由CRE的结构变异驱动,并进一步影响基因表达。此外,人工基因编辑造成的CRE改变,可以重塑cistrome,并进一步影响基因表达和性状差异。
0 5 其他因素
文章还提到实验的影响,这一部分很重要和实用。植物中缺乏高质量原生TF抗体,转基因标签是植物cistrome mapping的常用解决方案,但也可能制造非天然结合位点。另外,计算分析也会影响,文章中提到不同分析流程会得到不同cistrome。
4 如何判断一个结合位点是否有功能
前面提到过不是所有检测到的TF结合位点都有功能。如何判断一个结合位点是否有功能呢?文章中建议多组学联合。可以结合以下技术策略:
A.RNA-seq:TF结合附近基因是否表达改变;
B.TF突变体RNA-seq:改变是否依赖该TF;
C.组蛋白修饰ChIP-seq/CUT&Tag:是否具有活跃或抑制性表观标记;
D.ATAC-seq:结合位点是否位于开放染色质,对于DAP-seq获得的体外结合数据尤其有价值;
E.Hi-C/HiChIP/Hi-TrAC:远端CRE是否空间接触目标基因。
cistrome mapping只能告诉我们TF在哪里结合,要知道它是否调控基因表达,必须结合转录组基因表达数据、染色质可及性、组蛋白修饰标记和3D基因组数据。
5 展望未来
尽管植物的cistrome图谱构建仍落后于其他物种,但在整个植物科学界的共同努力下已取得显著进展。如前所述,针对转录因子的可用抗体稀缺性不太可能得到改善------尤其是在可获得基因组数量迅速增加、研究对象植物物种多样性不断扩大的背景下。因此,作者认为结合DAP-seq、RNA-seq、 ATAC -seq及三维基因组图谱技术,在体外开展大规模且高质量的顺式作用域图谱构建,仍是研究多种植物顺式作用域的重要策略。
作者也提到了单细胞测序,目前大多数cistrome数据来自bulk组织,实际上是许多细胞类型的混合结果。未来需要scATAC-seq构建细胞类型特异ACR图谱,scRNA-seq推断TF活性,
snCUT&Tag/scCUT&Tag直接看单细胞cistrome。但植物中单细胞TF cistrome mapping仍然很难,因为单细胞染色质量少,TF结合信号弱,背景噪声高,植物组织解离复杂。
总 结
植物转录因子的结合图谱并不是一个固定、容易测到的东西。它受到细胞类型、发育阶段、环境、染色质状态、辅因子、顺式元件结构以及实验分析流程的共同影响。我们测到的TF结合peak只是native cistrome的一个实验投影。解释这些peaks时,必须考虑TF剂量、辅因子、染色质、顺式元件结构以及实验/计算偏差。植物cistrome研究没有唯一最佳方法,而是需要根据研究问题,在ChIP-seq、CUT&RUN、CUT&Tag、DAP-seq、ATAC-seq等多种技术之间做取舍,并结合多组学数据解释其功能意义。未来植物cistromics的关键,是把ChIP/CUT/DAP等mapping技术与RNA-seq、ATAC-seq、单细胞组学和3D基因组数据整合起来,真正解析TF如何调控植物发育、胁迫响应和性状变异。
爱基百客深耕表观领域多年,旗下的ChIP-seq、CUT&Tag、DAP-seq、ATAC-seq等技术已协助多个植物cistrome研究发表高水平期刊。如您有相关技术需求,欢迎咨询~
爱基百客植物转录因子项目文章(部分)