Reciprocal conversion between annual and polycarpic perennial flowering behavior in the Brassicaceae
在十字花科植物中年生和多次开花多年生开花行为的互相转化

亮点
-
喜马拉雅须弥芥 和 内华达糖芥 是两个多年生植物模型
-
MADS-box 基因的剂量效应决定了一年生、二年生和多年生的生长习性
-
单个 MADS-box 基因就足以将一年生的开花行为转化为多年生
摘要
多年生作物的发展对于可持续农业和粮食安全具有重要潜力。然而,年生性和多年生性之间转化的进化过程尚不清楚。在此,我们利用两种十字花科植物,喜马拉雅须弥芥 和 内华达糖芥,作为多次开花多年生模型,揭示了多次开花多年生植物向二年生和一年生开花行为的转变是由三个密切相关的 MADS-box 基因的剂量决定的连续过程。这些基因表达模式、功能强度和组合的多样化使物种具有采用多种生活史策略的潜力。值得注意的是,我们发现这三个基因中的单个基因就足以将冬一年生或一年生的十字花科植物转变为多次开花的多年生植物。我们的工作阐明了植物多样生活史策略进化的遗传基础,并为未来多样化多年生十字花科作物的生成奠定了基础。
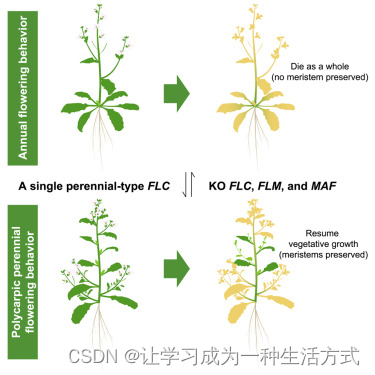
介绍
植物展示了各种各样的生活史策略。一年生植物在一年内完成其生命周期,大多数分生组织先发育成营养器官,然后在花诱导后变为生殖器官。开花后,整个植物经历衰老并最终死亡。二年生植物在第一年保持营养状态,第二年繁殖后死亡。基于繁殖次数(即植物繁殖的次数),多年生植物可以分为单次开花多年生和多次开花多年生。单次开花多年生植物有一个长的幼年期,可能持续数年,在此期间分生组织保持营养状态,对花诱导信号无反应。植物在成年后开花,开花也伴随着整个植物的衰老和死亡。另一方面,多次开花多年生植物也可能有一个延长的幼年期,但只有一部分分生组织在花诱导时进行花的转变,而其他分生组织保持营养状态。
多年生性和一年生性之间的进化转变是被子植物中最常见的进化转变之一。虽然一年生植物占大约10%的被子植物,但它们在不同科中的分布并不均匀。使用系统发育方法的祖先状态重建表明,一年生植物多次从多年生祖先进化而来。同属植物中一年生植物和多次开花多年生植物的共存表明这两种生活史策略之间的遗传差异并不大。然而,这一转变的进化模式和遗传基础仍然不太清楚。
十字花科中一年生模式物种拟南芥的许多近亲是多次开花多年生植物。其中,高山南芥(Arabis alpina)已成为理解多次开花多年生生长习性遗传基础的宝贵模式物种。高山南芥的一个一年生姊妹物种的引入促进了年生植物和多次开花多年生植物之间保守性和差异性的进一步分析。特别是,高山南芥在模拟冬季条件的长时间冷暴露后才开花,这一过程称为春化。最近的研究表明,关键花抑制基因FLOWERING LOCUS C(FLC)在春化途径中的差异表达对于十字花科中多次开花向单次开花的转变至关重要。拟南芥的FLC及其在高山南芥中的同源基因PERPETUAL FLOWERING1(PEP1)抑制开花,并在冷处理期间转录受到抑制。在拟南芥中,FLC的沉默是稳定的,使植物开花并最终经历全局停止(即单次开花)。相比之下,高山南芥中PEP1的抑制是暂时的,其转录在冷暴露后重新激活,从而限制了开花时间,并保留了营养腋芽和侧枝。在重新暴露于冷环境后,植物再次开花(即多次开花)。
需要注意的是,单独损伤PEP1并不会导致多年生性的丧失,因为高山南芥pep1突变体仍保持长寿(即持续开花)。因此,目前尚不清楚一年生植物是如何从多年生植物进化而来的,以及在十字花科中长寿性是如何调节的。此外,在同一植物属内多样生活史策略的进化方式,以及这些策略在不同物种中是否存在趋同进化,尚不清楚。为了回答这些问题,我们建立了须弥芥属和糖芥属属作为模型,以研究十字花科中多次开花多年生的共同遗传基础。具体来说,我们利用喜马拉雅须弥芥和内华达糖芥这两种多次开花多年生植物,以及相应的弱多次开花多年生/二年生植物卵叶须弥芥和一年生植物小花糖芥的自然变异来进行研究。
结果
本研究中不同生活史策略的定义
植物的生活史策略在生长室和自然条件下有所不同。在本研究中,个体植物的生活史策略是在生长室条件下进行评估和表征的。根据春化需求,植物可以分为两类:适应性春化和强制性春化。适应性春化响应的植物(以下简称适应性植物)不需要冷暴露即可开花,但在春化后会更早开花。相比之下,强制性春化需求的植物(以下简称强制性植物)如果没有经历冷温暴露则无法开花。考虑到自然生长季节的长度,我们将生长室条件下5个月内不开花的植物定义为本研究中的强制性植物。
需要注意的是,多次开花的生长习性不一定与强制性春化需求相关。适应性植物可以在没有冷处理的情况下开花并表现出多次开花的表型。因此,我们将本研究中观察到的多样生活史策略分为四个简单类别:一年生植物、冬一年生植物(具有适应性春化需求的单次开花植物)、二年生植物(具有强制性春化需求的单次开花植物)和多次开花多年生植物(具有适应性或强制性春化需求的多次开花植物)。
喜马拉雅须弥芥和内华达糖芥作为多次开花多年生模型
尽管通过胚胎救援方案成功生成了A. montbretiana和高山南芥的F1杂交种,但由于其模糊的生长习性(持续开花)和无法产生可行种子,进一步识别这些物种之间生活史策略差异的基因受到阻碍。为了解决这一问题,我们尝试在十字花科内建立其他多次开花多年生模型。在选择这些模型时,我们要求植物是二倍体、基因组小,并且有兼容的一年生、冬一年生或二年生植物。为此,我们首先筛选了十字花科中的所有属,发现须弥芥属和糖芥属植物在生活史性状上表现出丰富的多样性。
我们获得了两个二倍体多次开花多年生物种,分别是从仙台拟南芥种子库存中心(SASSC)获得的喜马拉雅须弥芥和从美国农业部农业研究服务局种质资源信息网络(GRIN)获得的内华达糖芥。喜马拉雅须弥芥生长在青藏高原,代表了一种研究极端环境中物种形成和生态适应的新模型系统,而内华达糖芥则是内华达山脉峰顶的特有种。我们在生长室条件下表征了这两个物种的生长习性。种子发芽后,喜马拉雅须弥芥开始产生莲座叶,并在长日照条件下持续生长时不开花。尽管一些植物在低温(4°C)暴露16周后开花,但需要20周的春化才能完全饱和开花反应,所有植物抽薹并产生完全发育的花序。所有花茎衰老后,新的腋芽在主茎和侧茎的节上开始出现。这些芽以营养状态生长并保持存活。

(A--I) 卵叶须弥芥 (A--C)、喜马拉雅须弥芥 (D--F)及其 F1 后代 (G--I) 的生长习性。植物在长日照条件下生长,照片分别拍摄于萌发后 18 周 (18W) (A, D, 和 G) 以及春化后 1 周 (AV1W) (B)、2 周 (AV2W) (H)、5 周 (AV5W) (E) 或 34 周 (AV34W) (C, F, 和 I)。请注意,所有植物都可以恢复营养生长(即形成新的营养芽)。
(J--Q) 小花糖芥 (J 和 K)、内华达糖芥 (L--N) 及其 F1 后代 (O--Q) 的生长习性。植物在长日照条件下生长,照片分别拍摄于萌发后 5 周 (5W) (J、L 和 O) 或 22 周 (22W) (K),以及春化后 3 周 (AV3W) (M)、4 周 (AV4W) (H) 或 32 周 (AV32W) (N 和 Q)。
(R) 卵叶须弥芥 (C. wal)、喜马拉雅须弥芥 (C. him) 及其 F1 后代的开花时间。植物开始开花时计算叶片总数。数据以小提琴图显示中位数(虚线)和四分位数(虚线),n ≥ 12。字母表示通过单因素方差分析(ANOVA)确定的显著差异,p < 0.01。植物在长日照条件下生长。NF,未开花。
(S) 卵叶须弥芥、喜马拉雅须弥芥 及其 F1 后代的春化敏感性。植物首先在长日照条件下生长 8 周,然后进行指定时间的春化。计算每次实验的开花植物百分比。每列顶部给出检查的植物数量。
(T) 卵叶须弥芥、喜马拉雅须弥芥 及其 F1 后代恢复营养生长的植物百分比。植物首先在长日照条件下生长 8 周,然后进行 22 周的春化,再返回室温。每隔 2 周计算一次恢复营养生长的植物数量,如 x 轴所示。误差条表示标准差 (SD)。
(U) 小花糖芥 (E. che)、内华达糖芥 (E. nev) 及其 F1 后代的开花时间。植物开始开花时计算叶片总数。数据以小提琴图显示中位数(虚线)和四分位数(虚线),n ≥ 11。字母表示通过单因素方差分析(ANOVA)确定的显著差异,p < 0.01。植物在长日照条件下生长。
(V) 内华达糖芥 及其与 小花糖芥 杂交产生的 F1 植物的春化敏感性。植物首先在长日照条件下生长 8 周,然后进行 x 轴上指定时间的春化。
(W) 内华达糖芥 及其与 小花糖芥 杂交产生的 F1 植物恢复营养生长的植物百分比。植物首先在长日照条件下生长 8 周,并春化 12 周。春化后 18 周计算恢复营养生长的植物数量。误差条表示标准差 (SD)。
比例尺:5.0 厘米 (A)--(Q)。
内华达糖芥 在长日照条件下表现出与 喜马拉雅须弥芥 相似的开花表型(图 1L--1N、1U--1W 和 S1A)。与 喜马拉雅须弥芥 相比,内华达糖芥 的完全春化需求仅为 10 周(图 1V)。此外,类似于高山南芥,内华达糖芥 在春化前或春化期间形成的所有腋芽都转变为开花,而春化后形成的芽则保持营养状态(图 1N)。再经过2个月的冷处理可以启动新一轮的开花(图 S1B 和 S1C)。因此,在我们的生长室条件下,喜马拉雅须弥芥 和 内华达糖芥 都是典型的多次开花多年生植物。
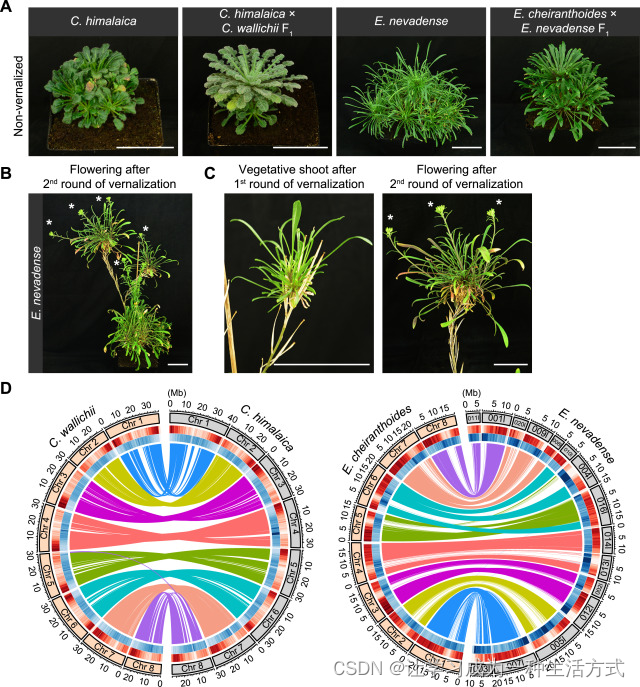
(A) 植物表型。这些植物在长日照条件下生长了1年。图中显示了未开花的F1植物。 (B) 内华达糖芥 的第二轮开花。内华达糖芥 植物再经过12周的春化处理后恢复到室温。请注意,这些植物可以启动第二轮开花。白色星号表示花序。 (C) 第一次春化后形成的营养芽和第二次春化后形成的花序(白色星号)。这株植物与(B)中的是同一株。 (D) 喜马拉雅须弥芥、卵叶须弥芥、内华达糖芥 和 小花糖芥 基因组重要特征的Circos显示。圆圈(从外到内)分别表示染色体或scaffold名称及大小(每5 Mb有刻度标记)、基因密度、转座元件密度以及同线性基因之间的链接。基因组特征是使用50 kb无重叠窗口计算的。红色和蓝色分别表示高密度和低密度。
比例尺:5.0厘米 (A)--(C)。
为了进行后续的遗传研究,我们寻找可以与 喜马拉雅须弥芥 和 内华达糖芥 进行杂交的二年生或一年生近缘种。对于 喜马拉雅须弥芥,我们从 SASSC 获得了 卵叶须弥芥。这种物种的原产地范围从伊朗到中亚和喜马拉雅山脉。卵叶须弥芥 在生活史策略上表现出高度的可塑性。在长日照条件下生长时,卵叶须弥芥 显示出弱多次开花多年生或冬一年生的生长习性(图 1A--1C)。尽管春化加速了开花,但大多数植物在长日照条件下以 22°C 产生60片叶子后即可开花(图 1R)。值得注意的是,虽然大多数植物在开花后死亡,但我们偶尔观察到无论是春化还是未春化的植物都能恢复营养生长(<10% 的植物,图 1C 和 1T)。这一结果表明,卵叶须弥芥 代表了多次开花多年生和冬一年生生长习性之间的过渡状态。
为了获得与 内华达糖芥 兼容的一年生植物,我们从 GRIN 筛选了106种 糖芥属 物种(参见 STAR 方法)并选择了 小花糖芥,也称为糖芥或除虫草壁花,这是一种短命的一年生植物,最近被用作研究十字花科中祖先和新型防御机制进化的模型。在长日照条件下生长时,小花糖芥 表现出类似于拟南芥的一年生生长习性。植物在开花前产生大约30片叶子,最终经历衰老并死亡(图 1J、1K 和 1U)。
在生长室的长日照条件下,喜马拉雅须弥芥 × 卵叶须弥芥 和 内华达糖芥 × 小花糖芥 的 F1 植物分别表现出与 喜马拉雅须弥芥 和 内华达糖芥 相同的多年生开花表型(图 1G--1I、1O--1Q、1T、1V 和 1W)。开花后,主茎节点处明显出现了营养枝(图 1I 和 1Q)。这些发现表明,多次开花多年生开花行为是一种显性性状,暗示十字花科中的一年生植物可能通过基因突变从其多年生祖先进化而来。综上所述,我们建立了两个研究十字花科中多次开花多年生开花行为的模型。拥有具有不同生活史性状的两属植物使我们能够更全面地理解这一科中生活史策略的进化模式。
与春化和多次开花多年生开花相关基因的遗传定位
我们对 喜马拉雅须弥芥、卵叶须弥芥、小花糖芥 和 内华达糖芥 的基因组进行了测序和组装(图 S1D;表 S1;参见 STAR 方法),并进行了基因定位,以识别两属中自然变异的开花行为这一生活史核心组成部分的相关基因。由于 内华达糖芥 和 小花糖芥 之间关系较远以及 内华达糖芥 的自交不亲和性,它们的 F1 杂交种几乎不育。为了克服这一困难,我们通过将 F1 植物与 小花糖芥 回交生成了 BC1F1 群体。对该群体中的188个个体进行基因组重测序分析成功识别了3个与春化需求显著相关的染色体区间(图 S2A)。通过注释拟南芥中的功能同源基因,我们获得了这些区间内的编码基因列表(表 S1)。正如预期的那样,在染色体6上的区间内发现了一个与 FLC 同源的基因。具体来说,我们在一年生亲本 小花糖芥 的 FLC 基因(简称 EcFLC)的第五密码子位置识别到一个导致翻译提前终止的失功能突变(图 S3C)。有趣的是,我们注意到其他两个区间中包含编码 FLC 类 MADS-box 转录因子的基因位点,即染色体2上的 FLOWERING LOCUS M(FLM)同源基因和染色体8上的 MADS AFFECTING FLOWERING(MAF)同源基因。接下来,我们基于营养生长恢复表型进行了遗传定位。显著的是,包含 FLC 和 FLM 基因的区间也与这一性状显著相关,其中 FLC 起主导作用(图 S2B)。
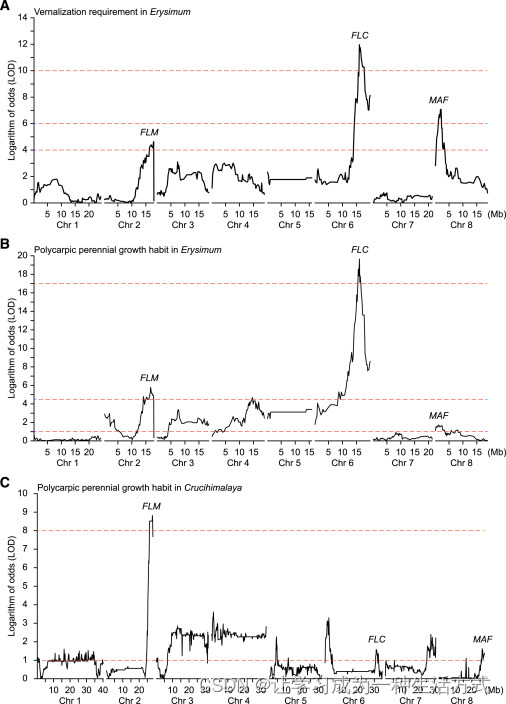
(A) 基于 须弥芥属 春化需求的遗传定位结果。 (B) 基于 须弥芥属 多次开花多年生生长习性的遗传定位结果。 (C) 基于 糖芥属 多次开花多年生生长习性的遗传定位结果。
图中展示了八条染色体(Chr1--8)。
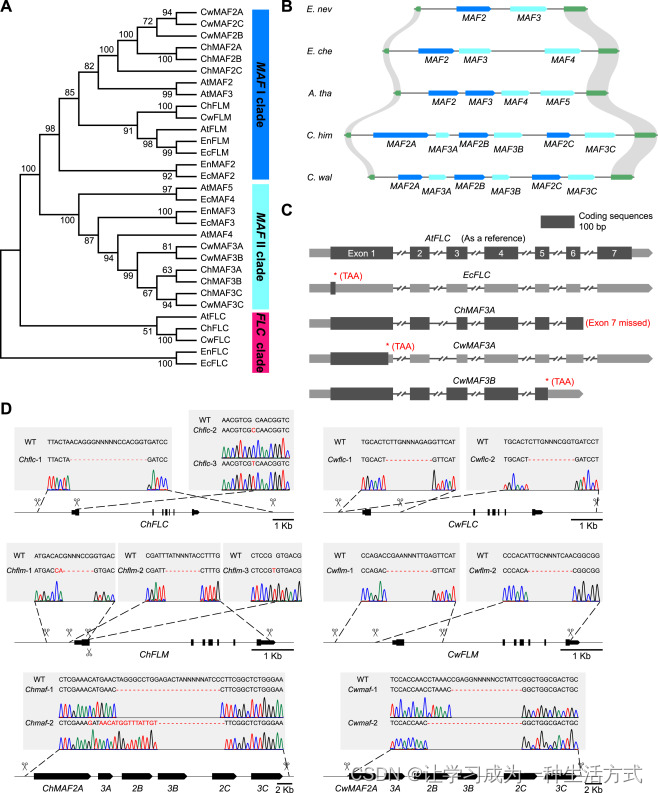
(A) 拟南芥、须弥芥属 和 糖芥属 中三种 FLC 类 MADS-box 基因的系统发育树。展示了三个进化支。拟南芥基因的登录号分别为 FLC (AT5G10140.1)、FLM (AT1G77080.8)、MAF2 (AT5G65050.3)、MAF3 (AT5G65060.1)、MAF4 (AT5G65070.3) 和 MAF5 (AT5G65080.2)。
(B) 拟南芥、须弥芥属 和 糖芥属 中 MAF 基因位点的基因组结构。同线性区域用阴影表示。
(C) 拟南芥、小花糖芥、喜马拉雅须弥芥 和 卵叶须弥芥 中 FLC 和 MAF 基因的基因结构。拟南芥中的 FLC 基因 (AtFLC) 作为参考。给出了突变的位置和性质。
(D) 通过 CRISPR-Cas9 技术在 须弥芥属 中生成 FLC 类 MADS-box 基因突变体。展示了 sgRNA 的位置(剪刀符号表示)、FLC 类 MADS-box 基因的基因组结构以及桑格测序结果。为创建大规模 DNA 缺失,每个基因设计了两个 sgRNA。WT,野生型。删除的核苷酸用红色虚线表示,而插入或替换的核苷酸用红色突出显示。
对于 须弥芥属 属,我们在 喜马拉雅须弥芥 和 卵叶须弥芥 之间生成了一个 F2 群体。对该群体的 250 个 F2 个体的分析确定了一个特定区间与多次开花多年生生长习性之间的强关联。值得注意的是,该区间与在 糖芥属 群体中染色体2上识别的区间具有基因组共线性,这两个区间都包含 FLM 基因(图 S2C)。此外,我们还检测到包含 FLC 和 MAF 基因的区间与多次开花多年生生长习性之间的弱关联(图 S2C)。这一结果可能归因于在我们的生长室条件下,卵叶须弥芥 表现出多次开花多年生/冬一年生的生长习性,与 喜马拉雅须弥芥 并无显著差异(见下文)。出于类似原因,我们没有在该群体中进一步进行春化需求的遗传定位。
在拟南芥中,FLC 属于一个花抑制基因的进化支,包括 FLM、MAF2、MAF3、MAF4 和 MAF5(图 S3A),其中 MAF2 至 MAF5 串联排列。同样,喜马拉雅须弥芥 和 卵叶须弥芥 中的 MAF 位点包含六个 MAF 基因,而 内华达糖芥 中包含两个 MAF 基因,小花糖芥 中包含三个 MAF 基因(图 S3B)。喜马拉雅须弥芥 中的 MAF3A 和 卵叶须弥芥 中的 MAF3A 和 MAF3B 基因可能因编码区内的提前终止密码子或截断而功能失活(图 S3C)。已显示 FLM 在环境温度范围内调节开花,高温通过选择性剪接结合无义介导的 mRNA 降解下调 FLM 表达。相比之下,MAF2--4 在控制开花时间方面与 FLC 功能重叠,但程度较低。因此,功能相似性和我们在两个不同属中的定位结果强烈表明,三种 FLC 类基因在十字花科中冗余地贡献于多次开花多年生生长习性。
FLC 类 MADS-box 基因冗余控制 须弥芥属 中的多次开花多年生开花
鉴于多次开花多年生生长习性是显性性状,我们通过使用成簇的规律间隔短回文重复序列(CRISPR)-Cas9 技术和农杆菌介导的转化生成 FLC、FLM 和 MAF 基因的失功能突变体来验证遗传定位结果。不幸的是,内华达糖芥 目前不适用于转化。为了生成缺失等位基因,我们通过在编码区内或其侧翼设计两个单导 RNA(sgRNA)来创建大片段缺失的突变体。由于目前尚不清楚六个 MAF 基因中的哪一个参与了多次开花多年生开花,我们删除了 须弥芥属 中的整个 MAF 位点。我们在 喜马拉雅须弥芥 中获得了 FLC、FLM 和 MAF 的单突变体,分别命名为 Chflc、Chflm 和 Chmaf。如图 2A 和 2C 所示,这三个突变体都具有开花的强制春化需求,这表明功能冗余。然而,这些突变体的饱和春化需求有所不同(图 2E)。与 Chflc 和 Chflm 相比,Chmaf 需要更长的冷处理(18 周)才能完全饱和春化需求。此外,我们注意到 ChFLC 或 ChFLM 的缺失导致植物在春化后恢复营养生长的能力下降(图 2D)。相比之下,Chmaf 单突变体表现出与野生型相同的多年生生长习性。因此,这些单突变体可以被认为是多次开花多年生或多次开花多年生/二年生开花植物。

(A) 喜马拉雅须弥芥 突变体的表型。拍摄照片时植物的年龄已给出。请注意,Chflc-1、Chflm-2 和 Chmaf-1 突变体在春化后可以恢复营养生长(即形成新的营养芽,插图中显示)。Chflc-1 Chflm-2 突变体表现出冬一年生的生长习性,它们在没有春化的情况下持续开花。
(B) Chflc-1 Chmaf-1 和 Chflc-1 Chflm-2 Chmaf-1 突变体的表型。
(C) 喜马拉雅须弥芥 突变体的开花时间。数据以小提琴图显示中位数(虚线)和四分位数(虚线),n ≥ 14。字母表示通过单因素方差分析(ANOVA)确定的显著差异,p < 0.01。
(D) 恢复营养生长的野生型和突变型 喜马拉雅须弥芥 (C. him) 植物的百分比。x 轴上每隔 2 周计算一次恢复营养生长的植物数量。误差条表示标准差(SD)。
(E) 喜马拉雅须弥芥 突变体的春化敏感性。植物首先在长日照条件下生长 8 周,然后进行指定时间的春化。完全饱和春化需求所需的时间用黑框表示。
(F) 卵叶须弥芥 (C. wal) 突变体的表型。拍摄照片时植物的年龄已给出。
(G) 野生型和突变型 卵叶须弥芥 植物的开花时间。植物开始开花时计算叶片总数。数据以小提琴图显示中位数(虚线)和四分位数(虚线),n ≥ 15。字母表示通过单因素方差分析(ANOVA)确定的显著差异,p < 0.01。
比例尺:图 (A)、(B) 和 (F) 中为 5.0 厘米。生长条件见 STAR 方法。
我们通过杂交生成了双突变体和三突变体。Chflc Chflm 双突变体表现出冬一年生的表型。这些植物在长日照条件下产生 150 片叶子后开花(图 2C),春化可以加速开花(图 2E)。在长日照条件下持续生长时,Chflc Chflm 植物最终可以抽薹并持续开花(图 2A),类似于高山南芥 pep1 突变体。无论是春化还是未春化的植物,营养生长的恢复都未发生(图 2A 和 2D)。有趣的是,Chflc Chmaf 双突变体和 Chflc Chflm Chmaf 三突变体都表现出一年生植物的特征(图 2B 和 2C)。特别是,Chflc Chflm Chmaf 三突变体在产生仅 10 片叶子后开花,最终衰老并死亡(图 2B 和 2C)。这些发现与遗传定位结果一致,并表明:(1) ChFLC、ChFLM 和 ChMAF 在春化调控中具有功能冗余;(2) ChFLC 和 ChFLM 的联合作用确保了 喜马拉雅须弥芥 中稳健的多次开花多年生生长习性;(3) ChMAF 强烈抑制开花,主要在春化中发挥作用。
与 Chflc 突变体不同,卵叶须弥芥 flc 突变体(以下简称 Cwflc)无需春化即可开花,并表现出与 Chflc Chmaf 双突变体类似的一年生生长习性(图 2A、2F 和 2G)。在 Cwflc Cwmaf 背景下进一步突变 CwFLM 导致更早的开花,但程度较轻(图 2G)。此外,Cwflc Cwflm Cwmaf 三突变体表现出比 Cwflc Cwflm 双突变体更显著的早开花表型(图 2F 和 2G)。这些结果表明,FLM 和 MAF 基因在 卵叶须弥芥 中的活性比在 喜马拉雅须弥芥 中弱,这可能解释了为什么 卵叶须弥芥 表现出不一致的多年生生长习性,并且经常表现为冬一年生植物(图 1T)。
综上所述,突变体表型分析揭示了 须弥芥属 中三个 FLC 类 MADS-box 基因在春化和多次开花多年生开花中的功能冗余。显著的是,通过同时使这三个基因失活,我们能够将多次开花多年生植物转化为类似一年生的植物。特别值得关注的是,不同组合的高阶突变体中观察到的多样生长习性进一步暗示,从多次开花多年生到一年生开花行为的进化是由这三个基因的剂量决定的连续过程。
三个 MADS-box 基因的剂量导致了 糖芥属 中生长习性的分化
为了进一步验证上述假设,我们在 小花糖芥 背景中构建了携带 内华达糖芥 FLC、FLM 和 MAF 的近等基因系(NIL),分别命名为 NIL-EnFLC、NIL-EnFLM 和 NIL-EnMAF(图 S4A)。我们通过杂交进一步生成了携带不同组合 内华达糖芥 MADS-box 基因的近等基因系。所有 NIL-EnFLC、NIL-EnFLM 和 NIL-EnMAF 系列在不同程度上表现出晚开花的表型(图 3A 和 3B),其中 EnFLC 对开花的抑制作用最大,其次是 EnMAF 和 EnFLM。特别是,NIL-EnFLC 表现出强制春化需求。有趣的是,结合 NIL-EnFLM 和 NIL-EnMAF 具有类似的效果。

(A) 示意图显示了八个近等基因系(NILs)。图中展示了八条染色体(纵向框架)。携带染色体1底部内华达糖芥片段的NIL-Ec被用作对照。
(B) NILs中EcSOC1和EcAP2的表达水平。请注意,这两个基因的表达模式与NILs的开花行为相关。
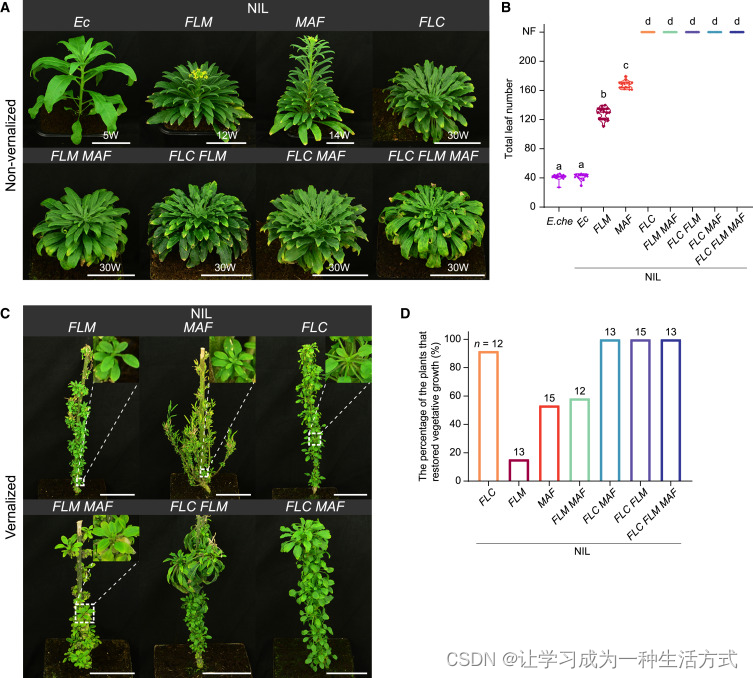
(A) 在长日照条件下生长的近等基因系(NILs)的表型。请注意,NIL-EnFLC、NIL-EnFLM EnMAF、NIL-EnFLC EnFLM、NIL-EnFLC EnMAF 和 NIL-EnFLC EnFLM EnMAF 在没有春化的情况下无法开花。携带染色体1底部内华达糖芥片段的NIL-Ec被用作对照。前缀"En"省略。
(B) 小花糖芥 (E. che) 和 NILs 的开花时间。数据以小提琴图显示中位数(虚线)和四分位数(虚线),n ≥ 12。字母表示通过单因素方差分析(ANOVA)确定的显著差异,p < 0.01。
(C) NILs 的表型。植物经过12周的春化处理。
(D) 在 NILs 中恢复营养生长的植物百分比。春化后18周计算恢复营养生长的植物数量。
比例尺:图 (A) 和 (C) 中为 5.0 厘米。生长条件见 STAR 方法。
携带单个 内华达糖芥 基因的近等基因系(NILs)表现出多样的生长习性。NIL-EnFLC 植物表现出多次开花多年生的开花表型(图 3C,S4B 和 S4C)。在春化后,NIL-EnFLM 和 NIL-EnMAF 植物也明显出现了营养芽(图 3C)。然而,这些芽的营养命运在长期内并不稳定,因为其中一些最终转变为生殖生长(图 3C,NIL-EnMAF 植物的插图),表明保持多次开花多年生开花行为的稳健性较低。这一表型在 NIL-EnMAF 植物中比在 NIL-EnFLM 植物中更为常见。因此,这两个 NILs 可以归类为多次开花多年生/冬一年生开花植物。值得注意的是,当 NIL-EnFLM 和 NIL-EnMAF 结合时,观察到营养生长的恢复(图 3C,S4B 和 S4C)。
综上所述,以上结果表明,与 须弥芥属 物种中的 FLC 相比,糖芥属 物种中的 FLC 在调节春化和多次开花中起主导作用。这一观察结果还可以解释为什么在 小花糖芥 × 内华达糖芥 BC1F1 群体中,包含 MAF 的基因组区间与多次开花多年生生长习性没有显著关联(图 S2C)。重要的是,这些结果进一步支持我们的结论,即三个 MADS-box 基因的剂量决定了一年生、二年生和多年生的开花行为。
三个 MADS-box 转录因子的共同下游靶点
为了进一步确认这三个 MADS-box 基因之间的功能冗余,我们尝试识别它们的下游靶基因。我们开发了针对 ChFLC 和 ChFLM 的特异性抗体。在 喜马拉雅须弥芥 中使用这些抗体进行染色质免疫沉淀测序(ChIP-seq)实验表明,ChFLC 和 ChFLM 共享一些作为直接靶点的开花时间和花图式基因(表 S2)。例如,我们发现这两个转录因子结合在 SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1 (ChSOC1)、SEPALLATA3 (ChSEP3)、APETALA3 (ChAP3)、SCHNARCHZAPFEN (ChSNZ)、GIBBERELLIN 3-OXIDASE2 (ChGA3OX2)、CONSTANS (ChCO)、CONSTANS-like 1 (ChCOL1) 和 SQUAMOSA PROMOTER BINDING PROTEIN-LIKE15 (Ch
SPL15) 的相同启动子区域(图 4A)。值得注意的是,所有这些基因也是高山南芥中 PEP1 的靶基因(表 S2),这表明多次开花多年生植物在十字花科中的下游调控网络具有进化保守性。与这些发现一致,在 Chflc Chflm Chmaf 三突变体中,与野生型相比,其靶基因 ChSOC1、ChSEP3 和 ChCO 的表达上调(图 4B)。
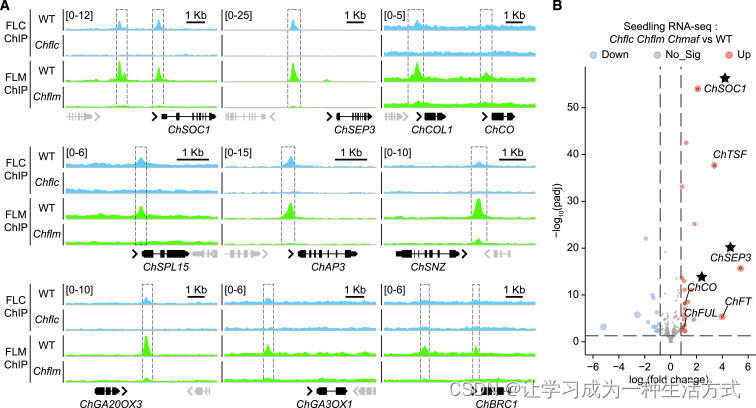
(A) 综合基因组查看器图显示了 ChFLC 和 ChFLM 的共同直接靶点。Chflc 突变体用作 ChFLC ChIP-seq 的对照,Chflm 用作 ChFLM 的对照。WT,野生型。每个基因的基因组区域(外显子和内含子分别用框和线表示)显示如下。靶基因以黑色显示,周围基因以灰色显示。方括号中的值表示标准化的读数计数。
(B) 火山图显示了野生型与 Chflc Chflm Chmaf 三突变体之间的差异表达基因。ChFLC 和 ChFLM 的靶基因用星号突出显示。
须弥芥属 中三个 MADS-box 基因的动态表观遗传和表达模式
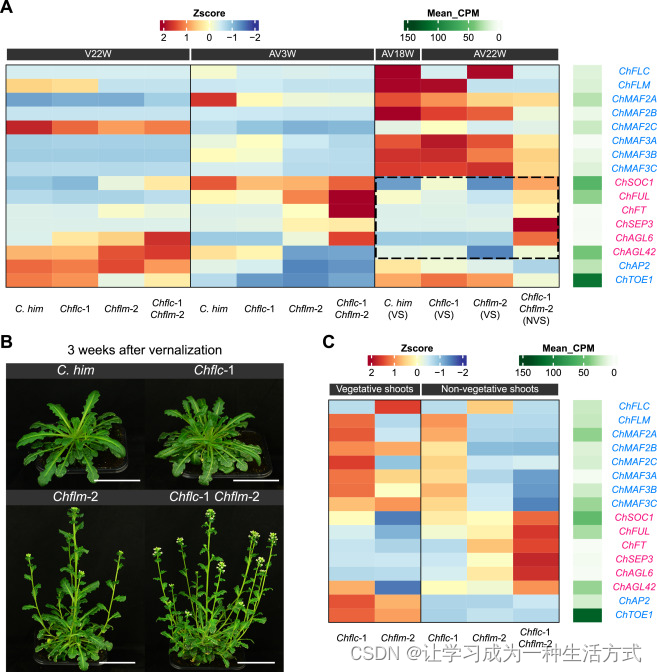
先前的研究表明,春化后 PEP1 的上调是恢复高山南芥营养生长和多次开花的关键步骤。然而,尚不清楚 FLC、FLM 和 MAF 在 喜马拉雅须弥芥 中是否也表现出类似的表达模式。为了研究这一点,我们进行了时间序列 RNA 测序(RNA-seq)实验。我们收集了春化前8周(8W)的莲座叶,春化12周和22周(V12W 和 V22W)的幼莲座叶,春化后3周(AV3W)的开花茎叶,以及春化后18周(AV18W)的营养芽叶。如图 5A 所示,ChFLC 在冷处理前(8W)表达水平很高。在 V12W 和 V22W,ChFLC 的转录水平大幅下降,在 AV3W 稍有增加,并在 AV18W 大部分恢复。这个表达模式在时间上与营养生长的恢复相关。

(A 和 B) 热图显示了 卵叶须弥芥 和 喜马拉雅须弥芥 (A) 及其 F1 植物 (B) 的基因表达模式。表达水平的变化由 RNA Z 得分显示,每个基因在所有时间点的绝对表达水平由 Mean_CPM 显示。
(C 和 D) 热图显示了 内华达糖芥 和 小花糖芥 (C) 及其 F1 植物 (D) 的基因表达模式。
(E) 喜马拉雅须弥芥 中 FLC 类 MADS-box 基因的 H3K27me3 ChIP-seq 轨迹。ChFLC 位点的核化区域由虚线框表示。方括号中的值表示 IP 样本的标准化读数/输入的标准化读数。
(F) F1 植物中 内华达糖芥(紫色)和 小花糖芥(洋红色)来源的 FLC 类 MADS-box 基因的 H3K27me3 ChIP-seq 轨迹。
在一年生拟南芥中,FLC 春化后抑制的稳定性与向 FLC 染色质中添加抑制性组蛋白修饰 H3K27me3 有关。相反,在多年生高山南芥中,PEP1 的短暂抑制主要归因于这种染色质修饰的调控和维持上的差异。因此,我们通过 ChIP-seq 实验在相同时间点的样本中检测了 ChFLC 位点的 H3K27me3 水平。8W 样本中的 H3K27me3 水平较低(图 5E)。在12周的冷暴露后,我们观察到 ChFLC 的外显子1和内含子1起始处 H3K27me3 水平增加,这在空间上与拟南芥 FLC 中的核化区域重叠。H3K27me3 的沉积在 V22W 时进一步增加并扩散至整个 ChFLC 位点,到 AV3W 时开始减少,在 AV18W 时已无法检测到(图 5E)。这些结果表明,ChFLC 表现出与其在高山南芥中的同源基因相同的行为。
ChFLM 的表达模式与 ChFLC 不同:其转录在 V12W 时没有显著减少,但在 V22W 时下降了约 40%(图 5A 和 S5A-S5C)。有趣的是,ChFLM 在 AV3W 时表达大幅减少,并在 AV18W 时随后上调。这些表达模式表明 ChFLM 受冷温抑制也是短暂的,但相对于 ChFLC 来说有所延迟。ChFLM 的 H3K27me3 动态与 ChFLC 类似。H3K27me3 在 V22W 时明显沉积,在外显子1和内含子1之间的连接区域达到最高水平(图 5E)。H3K27me3 水平在 AV3W 时较高,在 AV18W 时变得不可检测。

(A--C) 拟南芥 (A, FRISF2FLC)、须弥芥属 (B) 和 糖芥属 (C) 在 22°C 和 28°C 下 FLM 的可变剪接。在拟南芥中,FLM 受到温度依赖的可变剪接,产生两种剪接变体,FLM-β 和 FLM-δ。具体来说,高温通过可变剪接结合无义介导的 mRNA 降解抑制 FLM。然而,在 22°C 和 28°C 下,我们未在 须弥芥属 (B) 或 糖芥属 (C) 物种中观察到高水平的 FLM-δ 异构体,表明 FLM 的转录后调控可能没有进化保守性。y 轴表示以百万计数(BPM)显示的转录本丰度。展示了两种 FLM 异构体,FLM-β 和 FLM-δ。与可变剪接相关的区域用黄色突出显示,并在右侧放大。
(D 和 E) 小花糖芥 (D) 和 内华达糖芥 (E) 中 FLC 类 MADS-box 基因位点的 H3K27me3 ChIP-seq 轨迹。显示了基因组区域(外显子和内含子分别用框和线表示)。方括号中的值表示 IP 样本的标准化读数/输入的标准化读数。NV_5W 和 NV_8W,春化前5周或8周的植物;5W_V12W,5周大的植物春化12周;5W_AV3W,5周大的植物春化12周并恢复到室温3周。
ChMAF 基因的表达较为复杂。由于其极低的表达水平(图 5A)和编码截短蛋白(图 S3C),ChMAF3A 可能是非功能性的。ChMAF2C 的表达在 V12W 时升高,随后降低。ChMAF2A、ChMAF2B、ChMAF3B 和 ChMAF3C 表现出与 ChFLC 和 ChFLM 相似的表达模式。ChMAF2C 的 H3K27me3 动态与 ChFLM 类似,而 ChMAF2A、ChMAF2B、ChMAF3B 和 ChMAF3C 的模式与 ChFLC 类似(图 5A)。这些结果表明,ChMAF2A、ChMAF2B、ChMAF3B 和 ChMAF3C 可能参与了 喜马拉雅须弥芥 的多年生开花。
与三个 MADS-box 基因的动态表达模式一致,促进开花和花图式基因如 ChSOC1、FRUITFULL (ChFUL)、FLOWERING LOCUS T (ChFT)、ChSEP3、AGAMOUS-LIKE 6 (ChAGL6) 和 ChAGL42 表现出相反的模式(图 5A):在 8W、V12W 和 V22W 表达较低,在 AV3W 表达较高,并在 AV18W 表达在基础水平。相比之下,开花抑制基因如 ChAP2 和 TARGET OF EAT1 (ChTOE1) 在春化过程中下调,并在春化后新形成的营养芽中恢复表达(图 5A 和 S6)。
我们进一步调查了 喜马拉雅须弥芥 和 卵叶须弥芥 杂交后代 F1 植物的基因表达模式。与两亲本植物的 RNA-seq 分析结果相似,F1 代植物中的 FLC 类 MADS-box 基因在春化前后也表现出振荡的表达模式(图 5B)。在 AV18W 时,仅有两个基因,ChMAF2C 和 CwMAF2A,未恢复表达。因此,这些结果表明,喜马拉雅须弥芥 和 卵叶须弥芥 之间 FLC 类 MADS-box 基因的自然变异不太可能源于转录水平的差异。相反,在 卵叶须弥芥 中观察到的 FLM 和 MAF 基因功能减弱可能归因于蛋白质序列的变化。
我们对 内华达糖芥、小花糖芥 及其 F1 植物进行了类似的 RNA-seq 和 ChIP-seq 实验,并得到了类似的结果(图 5C、5D、S5D 和 S5E)。特别是,F1 代植物中的多年生 FLC 类 MADS-box 基因表现出与 内华达糖芥 相似的动态表达和 H3K27me3 沉积模式。同样,一年生 FLC 类 MADS-box 基因也保持了与 小花糖芥 相似的模式(图 5D)。这些结果表明,内华达糖芥 和 小花糖芥 之间 FLC 类 MADS-box 基因的功能分化主要由基因内顺式调控元件的变化决定。
综上所述,我们全面的 RNA-seq 和 H3K27me3 ChIP-seq 分析表明,除了 FLC 外,FLM 和 MAF 也在多年生和一年生植物中表现出不同的表达模式和表观遗传动态。重要的是,这些差异与 须弥芥属 和 糖芥属 中生活史策略的转变密切相关。
十字花科中与生活史相关的多样开花行为进化的统一假设
基于来自两个属的四个物种的上述发现,以及具有不同生长习性的突变体和近等基因系(NILs),我们提出了一个关于 FLC 相关 MADS-box 基因在十字花科中调控开花行为的统一假设(图 6)。重要的是,这一假设提供了一个合理的解释,即这三个基因在进化过程中的多样化如何促成了同一属内多样生活史策略的进化。这种多样化涉及表达模式(水平、动态和调控机制)的变化、表观遗传沉默的稳定性(通过春化稳定或不稳定地沉默)、春化反应和营养生长恢复的功能强度(强或弱)(图 6A)。这些变化共同导致了基因剂量依赖的开花时间和多次开花调控,从而导致植物生活史策略进化的连续过程。
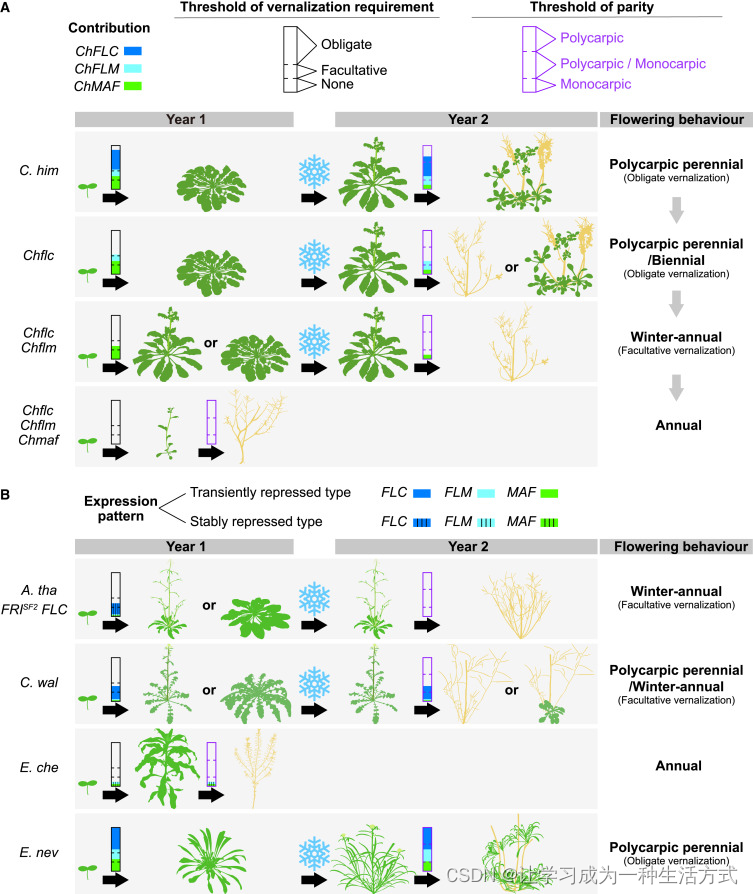
(A) 在 喜马拉雅须弥芥 中,生活史相关的开花行为调控。虽然 ChFLC、ChFLM 和 ChMAF 都在春化后暂时受到抑制,但它们对春化需求和生殖方式的贡献各不相同(用不同颜色表示;无颜色表示零贡献)。这三个基因的综合效应根据假定的春化需求阈值(虚黑线)(即强制性、适应性或无需求),以及生殖方式阈值(虚品红线)(即多次开花、多次/单次开花或单次开花),定义了开花行为。
(B) 在拟南芥、须弥芥属 和 糖芥属 中,生活史相关的开花行为调控。除了对春化需求和生殖方式作出不同贡献外(A),这些基因还表现出不同的表达模式(即是否通过春化稳定或暂时抑制)。每种物种或突变体的开花行为显示在右侧。为了简化,仅显示生命周期的前两年,绿色和黄色分别代表活体和死亡的植物部分。
由于一年生植物是从多次开花多年生植物衍生而来的,我们认为多次开花多年生是祖先状态。在诸如 喜马拉雅须弥芥 和 内华达糖芥 等植物中,所有三个 MADS-box 基因都调控春化反应和多次开花(图 6A 和 6B)。冷处理抑制的不稳定性导致这些基因在春化后表达恢复,从而促进营养生长和多次开花。需要注意的是,这三个基因的存在确保了稳健的多次开花多年生生长习性。任何一个基因突变的植物偶尔会表现出二年生特征(例如,图 6A 中的 Chflc 突变体)。此外,由于这三个基因功能强度不同,不同的单突变体可以表现出不同程度的多次开花多年生或二年生开花表型(图 2A-2C)。相比之下,在高山南芥中,PEP1 可能起着压倒性的主导作用。因此,PEP1 的单个突变导致春化需求消失和多次开花受损。虽然从一年生植物 A. montbretiana 引入 MAF 相关基因可以延迟 高山南芥 的开花,但 高山南芥 自身的 FLM 和 MAF 基因在多次开花多年生中的作用仍需进一步研究。事实上,pep1 突变体并未表现出一年生的生长习性,进一步表明 FLM 和 MAF 在 高山南芥 中与 PEP1 具有冗余作用。
三个 FLC 类 MADS-box 基因中的两个突变会导致冬一年生或一年生的开花行为。不同的突变组合可以导致不同的表型。例如,虽然 Chflc Chflm 双突变体表现出冬一年生的表型(图 6A),但 Chflc Chmaf 植物表现出一年生的生长习性(图 2B 和 2C)。尽管 FLC 类 MADS-box 基因的表达模式和调控机制在 喜马拉雅须弥芥 和 卵叶须弥芥 之间相似(图 5A),但 CwFLM 和 CwMAF 在 卵叶须弥芥 中的功能比在 喜马拉雅须弥芥 中弱(图 2F 和 2G)。因此,卵叶须弥芥 表现出多次开花多年生/冬一年生的生长习性(图 6B)。
这些三个基因的完全丧失或基因表达模式的改变会导致一致的一年生生长习性。在 喜马拉雅须弥芥 中,Chflc Chflm Chmaf 三突变体表现出一年生特征,植物在开花后最终衰老和死亡(图 6A)。在 卵叶须弥芥 中,由于 CwFLM 和 CwMAF 在春化和多年生开花中起次要作用,Cwflc 单突变体也表现出一年生开花表型。在 小花糖芥 中,EcFLC 突变(图 S3A)。EcFLM 和 EcMAF2 在春化后稳定受抑(图 5F,S5D 和 S5E),从而导致一年生生长习性(图 6B)。
在模式植物拟南芥中,MAF 基因在开花中的功能大大减弱,而 FLM 主要参与环境温度下的开花时间控制。值得注意的是,冷处理可以稳定地沉默 FLC、FLM 和 MAF。因此,拟南芥的生活史策略主要由 FLC 定义。具体而言,五种主要的 FLC 单倍型在功能上不同,在 FLC 表达水平和表观遗传沉默速度上有所不同,导致从二年生和冬一年生到一年生的生长习性的多样性。特别是,如 Col-0 等品种中 FLC 表达的促进因子 FRIGIDA 的突变导致了一年生的特征。
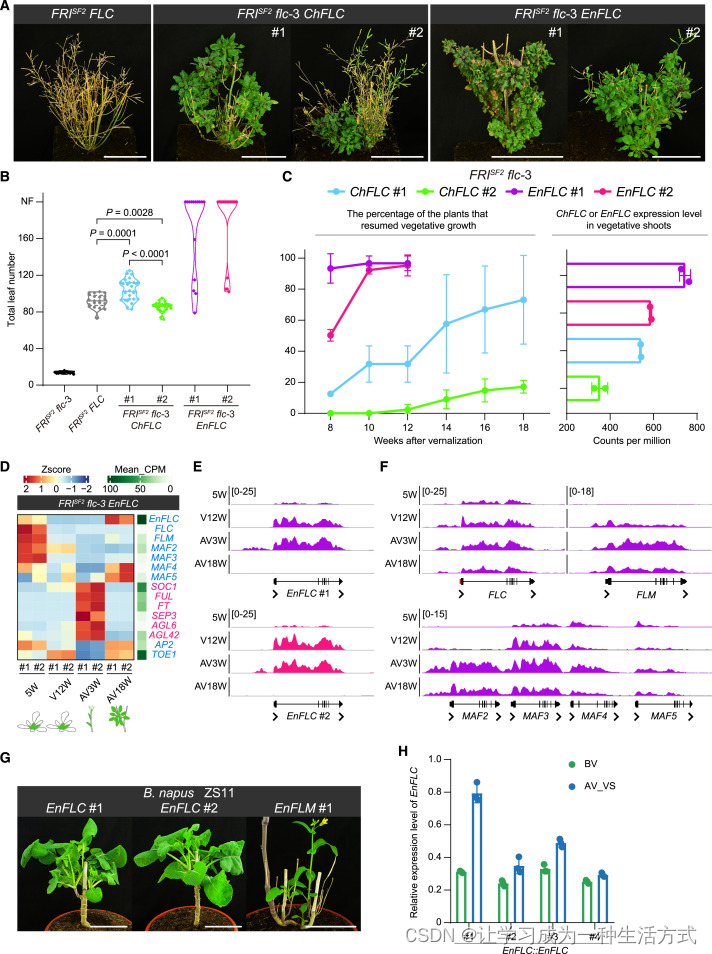
(A) 转基因拟南芥植物在 AV18W 时的表型。请注意,与 FRISF2 flc-3 和 FRISF2 FLC 相比,FRISF2 flc-3 ChFLC 和 FRISF2 flc-3 EnFLC 植物在春化后可以恢复营养生长。
(B) 开花时间测量。数据以小提琴图显示中位数(虚线)和四分位数(虚线),n ≥ 13。
(C) 恢复营养生长的转基因植物百分比。左侧显示了通过 RNA-seq 确定的 ChFLC 或 EnFLC 的表达水平。
(D) 热图显示了 FRISF2 flc-3 EnFLC 植物中 EnFLC 和拟南芥开花时间及花图式基因的表达模式。每条线进行了两个生物重复。
(E 和 F) FRISF2 flc-3 EnFLC 植物中 EnFLC (E) 和三个拟南芥 FLC 类 MADS-box 基因位点 (F) 的 H3K27me3 ChIP-seq 轨迹(线 #1 用洋红色表示,线 #2 用紫色表示)。
(G) 转基因油菜植物的表型。第1年形成的枯死茎已被移除。显示了 EnFLC 的两个独立品系(#1 和 #2)和 EnFLM 的一个品系。
(H) 转基因油菜植物春化前(BV)和春化后(AV) EnFLC 的表达水平。显示了四个独立品系(#1--#4)。春化后采集营养芽(AV_VS)。误差条表示标准差(n = 3)。
结果显示了两个独立品系(#1 和 #2)(A--E)。比例尺:图 (A) 中为 5.0 厘米,图 (E) 和 (F) 中为 10.0 厘米。生长条件见 STAR 方法。
将一年生或冬一年生植物转化为十字花科中多次开花多年生植物
基于上述假设,我们试图将一年生植物拟南芥和 小花糖芥 转化为多次开花多年生植物(即具有多年生开花行为的植物)。然而,目前尚不清楚春化后 FLC 的重置是否需要未知的上游调控因子。特别是,这些因子是否仅存在于多年生基因组中仍然未知。尽管如此,我们首先生成了转基因 小花糖芥,该转基因表达来自同一属的多次开花多年生植物 内华达糖芥 的 FLC、FLM 或 MAF 基因。所有基因均由其自身的 内华达糖芥 调控序列驱动(即 EnFLC::EnFLC、EnFLM::EnFLM 和 EnMAF2::EnMAF2)。在 T1 代中,我们观察到具有不同生长习性的转基因植物。具体来说,一些转基因系如 EnFLC::EnFLC #1、EnFLM::EnFLM #2 和 EnMAF2::EnMAF2 #1 不仅表现出强制春化需求(图 S7B),而且在春化后恢复营养生长(图 S7A 和 S7C)。RNA-seq 进一步揭示了这些植物中 EnFLC、EnFLM 和 EnMAF2 的表达水平约为表现出弱表型的系的四倍(图 S7D 和 S7E)。因此,这些结果验证了 糖芥属 近等基因系(NILs)的结果,并表明当表达水平超过某一阈值时,任何一个 内华达糖芥 的 FLC 类 MADS-box 基因都足以将一年生 小花糖芥 转化为多次开花多年生植物。

(A) 植物在春化后24周的表型。所有植物均春化12周后恢复到室温。比例尺,5.0厘米。
(B) 开花时间测量。植物开始开花时计算叶片总数。数据以小提琴图显示中位数(虚线)和四分位数(虚线),n ≥ 16。植物在长日照条件下生长。NF,未开花。
(C) 植物在春化后18周恢复营养生长的百分比。植物首先在长日照条件下生长,然后进行春化超过12周。
(D) 转基因植物中的基因表达水平。EnFLC、EnFLM 和 EnMAF2 的表达水平通过 RNA-seq 揭示(每百万计数)。误差条表示标准差(SD)。
(E) 热图显示转基因植物中开花时间和花图式基因的表达模式。样品在 AV18W 时收集。表达水平的变化通过 RNA Z 得分显示,每个基因在所有时间点的绝对表达水平由 Mean_CPM 显示。
(F) 热图显示 FRISF2 flc-3 ChFLC 植物中开花时间和花图式基因的表达模式。样品按照指示收集。
(G 和 H) FRISF2 flc-3 ChFLC 植物中 ChFLC (G) 和其他 MADS-box 基因位点 (H) 的 H3K27me3 ChIP-seq 轨迹。显示了基因组区域(外显子和内含子分别用框和线表示)。方括号中的值表示标准化读数。图 (G) 中显示了两个独立品系(#1 和 #2)的结果。
图 (B) 和 (D) 中检查的植物数量见表 S4。
我们在异种植物拟南芥中表达了 EnFLC 或 ChFLC,使用它们各自的调控序列(EnFLC::EnFLC 和 ChFLC::ChFLC)。为了在植物中实现 EnFLC 和 ChFLC 的高表达水平,我们转化了 FRISF2 flc-3 (Col-0) 植物(以下简称野生型)并为每个构建获得了超过 20 个 T1 转基因系。我们使用 FRISF2 FLC (Col-0) 植物作为对照。与野生型相比,FRISF2 FLC、EnFLC::EnFLC 和 ChFLC::ChFLC 植物在长日照条件下开花较晚(图 7B)。值得注意的是,大多数 EnFLC::EnFLC 植物(系 #1 和 #2)在 5 个月后仍未开花(图 7B)。这些植物暴露在低温环境 12 周后诱导开花。值得注意的是,与 FRISF2 FLC 系列相比,EnFLC::EnFLC 系列(#1 和 #2)在春化后可以恢复营养生长(图 7A 和 7C)。后来出现的腋芽以营养状态生长(图 7A)。
ChFLC::ChFLC 系列(#1 和 #2)的表型比 EnFLC::EnFLC 系列较轻。尽管 ChFLC::ChFLC 的开花时间与 FRISF2 FLC 相当(图 7B),但营养生长的恢复仍然明显(图 7A 和 7C)。与基因剂量效应对生长习性的关系一致,EnFLC::EnFLC 系列(#1 和 #2)中的 ChFLC/EnFLC 表达水平高于 ChFLC::ChFLC 系列(#1 和 #2)(图 7C)。因此,这些结果表明,较高水平的 ChFLC 或 EnFLC 足以将冬一年生的拟南芥转化为多次开花的多年生植物。
接下来,我们对多次开花多年生拟南芥植物进行了详细的分子表征。RNA-seq 实验表明,EnFLC 在 EnFLC::EnFLC 植物中春化后受到抑制,并且在新形成的营养腋芽的叶片中表达恢复到较高水平(图 7D)。另一方面,FLC (flc-3)、FLM、MAF2 和 MAF3 在春化后保持稳定抑制。有趣的是,尽管 MAF4 和 MAF5 的表达非常低,但这些基因的表达在春化后被重置。促进开花和花图式基因如 SOC1、FUL、FT 和 SEP3 的表达与 EnFLC 的表达趋势相反(图 7D)。
ChIP-seq 数据进一步表明,EnFLC 位点的 H3K27me3 水平在春化期间增加,并在植物恢复到室温后减少(图 7E)。相比之下,内源性 FLC (flc-3)、FLM、MAF2 和 MAF3 位点在春化后稳定沉默(图 7F)。在 ChFLC::ChFLC 系列中获得了类似的结果(图 S7F--S7H)。因此,这些发现支持了年生和多年生 FLC 基因之间表观遗传沉默差异是由 FLC 非编码区顺式多态性引起的这一观点。值得注意的是,我们的结果表明,负责在春化后重置 FLC 表达的上游调控因子仍存在于一年生的拟南芥中。
最后,我们尝试生成多次开花多年生的油菜植物。为了在植物中实现多年生 FLC 类基因的高表达水平,我们将这些基因转化到冬一年生品种 ZS11 中。在 25 个 EnFLC::EnFLC T1 系列中,有 9 个系列在春化后可以恢复营养生长。这从新的腋芽长出莲座叶且不伸长节间中得以显现(图 7G)。与这些发现一致,我们发现 EnFLC 表达在春化后被重置(图 7H)。需要注意的是,引入 EnFLM::EnFLM 不足以诱导多次开花多年生的开花行为,因为春化后形成的所有腋芽仍然进行生殖生长(图 7G)。这可能归因于异种植物中 EnFLM 表达不足或不正确。
综上所述,我们已成功将三种一年生或冬一年生十字花科植物转化为多次开花植物。值得注意的是,这些实验确认了基因剂量在生活史相关开花行为进化中的关键作用。观察到的不同表型可能归因于异种植物中 MADS-box 基因的差异表达水平。此外,我们的结果表明,拟南芥、小花糖芥 和油菜具有在生活史策略之间转换的遗传背景。值得注意的是,单个外源 FLC 或 MAF 基因的引入足以实现将一年生或冬一年生植物转化为多次开花多年生植物。
讨论
多年生植物必须保持至少一个分生组织保持不定性,以保留在第一个生长季后产生新器官的能力。我们的结果表明,由三个 FLC 类基因(即 FLC、FLM 和 MAF)控制的多年生开花行为对于在开花后保留营养分生组织至关重要,这是十字花科多年生性的关键特征。然而,需要强调的是,多年生开花仅是十字花科多年生性的先决条件。多年生性和一年生性应该被视为由许多其他相互作用的特征组成的综合症,如叶片和根的解剖结构、资源利用效率和源-汇动态的调控。尚不清楚这三个 FLC 类基因是否除了开花时间和多次开花外还调控其他多年生特征。由于 喜马拉雅须弥芥 flc flm maf 三突变体表现出与拟南芥相似的一年生生长习性,这表明这三个基因可能在控制植物寿命方面也起着冗余作用,尽管确切的机制尚待进一步研究。
尽管在温暖条件下恢复物种如高山南芥和日本白花芥中 FLC 同源基因表达的过程是确保十字花科中多次开花多年生开花的保守关键过程,但这种现象的具体机制仍不清楚。表达 ChFLC 或 EnFLC 的转基因拟南芥和 小花糖芥 植物表现出与 喜马拉雅须弥芥 和 内华达糖芥 相同的多次开花多年生生长习性(图 7 和 S6)。这些结果以及 FLC 编码序列的高度保守性支持了多年生和一年生 FLC 同源基因之间的主要差异在非编码区,可能是第一个内含子。需要注意的是,高山南芥 PEP1 在异种表达时并不能赋予拟南芥多次开花多年生生长习性。这种差异可能源于 PEP1 在拟南芥中的异常表达,未能达到某一阈值水平。
(A) 热图显示了野生型和不同突变背景下的基因表达模式。VSs,营养芽;NVSs,非营养芽。请注意,Chflc-1 和 Chflm-2 都是多次开花多年生/二年生开花植物,而 Chflc-1 Chflm-2 双突变体是冬一年生开花植物(图 2A-2C)。一致地,花促进基因在 Chflc-1 或 Chflm-2 单突变体的 VS 中保持抑制状态,但在 AV22W 时 Chflc-1 Chflm-2 双突变体的 NVS 中上调(虚线框)。植物春化 22 周(V22W)后恢复到室温 3 周(AV3W)、18 周(AV18W)和 22 周(AV22W)。
(B) 植物开花表型。所有植物春化 22 周后恢复到室温 3 周。请注意,Chflm-2 和 Chflc-1 Chflm-2 突变体比 Chflc-1 开花更早。比例尺,5.0 厘米。
(C) 热图显示了 Chflc-1、Chflm-2 和 Chflc-1 Chflm-2 突变体在 AV22W 时 VS 和 NVS 中的基因表达模式。请注意,Chflc-1 和 Chflm-2 单突变体在 AV22W 时的 NVS 中花促进基因如 ChSOC1 和 ChFUL 上调。
有趣的是,我们的研究结果表明,负责在春化后重置多年生 FLC 单倍型的上游调控因子在一年生植物如拟南芥和 小花糖芥 中依然存在。因此,考虑到拟南芥拥有丰富的遗传资源和高效的基因组编辑系统以及相对较短的生命周期,我们可以使用转基因拟南芥植物作为模型来阐明未来 FLC 不稳定表观遗传沉默和重置的分子机制。事实上,在拟南芥的开创性研究中已揭示,FLC 表达的变异受顺式定位的单核苷酸多态性控制。
过去十年中多年生作物研究最令人兴奋的前沿是新多年生作物物种的开发。例如,Zhang 等人最近报告通过将驯化的亚洲一年生水稻(Oryza sativa)与其多年生非洲亲缘种 Oryza longistaminata 杂交,成功培育出多年生水稻。或者,Yu 等人利用高质量的基因组组装来鉴定二倍体水稻中农艺重要基因的同源基因,并通过基因组编辑成功实现了 Oryza alta 各种性状的目标改良。因此,考虑到同一科内的低遗传差异以及成功将拟南芥、小花糖芥 和油菜转化为多次开花多年生植物,我们在 须弥芥属 和 糖芥属 中鉴定的涉及多年生到一年生转变的基因为未来其他十字花科多年生作物的发展铺平了道路。
研究的局限性
我们的研究有两个主要局限性。首先,驱动从多次开花多年生到一年生开花行为转变的 FLC 类 MADS-box 基因中突变的具体性质尚不清楚。这主要是由于我们多次开花多年生模型与其一年生亲缘种之间的巨大遗传距离。其次,尽管我们能够在生长室条件下恢复冬一年生油菜的营养生长,但尚不清楚这些植物在田间是否会表现为真正的多次开花多年生。此外,开花行为变化对产量的潜在影响目前尚不清楚。因此,需要进一步的遗传改良和大规模田间试验,以生成具有实际价值的多次开花多年生油菜品种。