Disco : Densely-overlapping Cell Instance Segmentation via Adjacency-aware Collaborative Coloring
ICLR 2026
Disco:基于邻接感知协同着色的密集重叠细胞实例分割方法
密集重叠细胞实例分割:该任务最大的行业痛点。病理切片里的癌细胞、免疫细胞经常高度聚集、边界严重交叠,甚至三个及以上细胞挤成一团,传统方法要么把多个细胞当成一个(欠分割),要么把一个细胞拆成好几个(过分割),漏检、错检率极高。
基于邻接感知协同着色:先看全局细胞的相邻关系,用着色规则先把大部分细胞分开,再把挤成一团的难分细胞单独拎出来精准拆解
着色:核心是图论里的「图着色规则」。简单来说,我们把每个细胞当成一个 "点",两个细胞挨在一起(空间相邻)就连一条 "线",这样整张病理图里的细胞,就组成了一张「细胞邻接图」。图着色的核心规则是:挨在一起的两个点(相邻细胞),绝对不能用同一个颜色。只要给每个细胞分对了颜色,就能从根源上把相邻的细胞彻底分开,不会出现混淆。
邻接感知:传统的细胞分割方法,只看单个细胞的边界、形状、像素灰度这些局部信息,而这个算法是全局感知所有细胞的相邻关系、拓扑结构,不是只看单个细胞,而是看 "这个细胞和谁挨着、和谁挤成了团",从全局规则上避免把相邻细胞搞混
协同着色:之前的图着色方法分两种极端:要么用 2 着色(只能处理两个细胞相邻的简单场景,三个细胞互相挨着就会失效),要么用 4 着色(四色定理,啥场景都能处理,但模型冗余度极高,AI 学习难度大、效果差)。
而 Disco 的协同着色,用了「分治协同」的思路:
- 先处理占绝大多数的简单区域:用 2 种颜色,把两个两个相邻的细胞(二分图结构)高效分开;
- 再攻坚少数复杂区域:把三个及以上互相挨着、2 着色搞不定的 "冲突细胞簇" 单独标记出来,用专属的 "冲突色" 标注;
- 最后用专门的算法机制,让 AI 在特征空间里把这些冲突的细胞也彻底分开;
- 简单区域和复杂区域两套机制协同工作,既避免了 2 着色的能力不足,也规避了 4 着色的冗余低效,实现了效率和精度的平衡。
摘要
精准的细胞实例分割是数字病理分析的基础。基于轮廓检测与距离映射的现有方法,在处理复杂、密集的细胞区域时仍面临巨大挑战。基于图着色的方法为此任务提供了新范式,但该范式在密集重叠、拓扑结构复杂的真实场景中的有效性尚未得到验证。
针对这一问题,我们发布了大规模数据集GBC‑FS 2025 ,其中包含高度复杂、密集的亚细胞排布。我们首次在四个不同数据集上系统分析了细胞邻接图的着色特性 ,并揭示一项重要发现:真实场景中的绝大多数细胞图均为非二分图,普遍存在大量奇环(以三角形为主)。这使得简单的二着色理论不足以处理复杂组织,而更高着色数的模型又会带来表征冗余与优化困难。
##注释
细胞邻接图的着色特性:把每个细胞 = 一个点 ;两个细胞挨在一起 = 连一条线 ;整张图就变成了细胞邻接图 ;着色 :给每个点上色,相邻点不能同色
这就是图论里的图着色,用来把挨在一起的细胞 "区分开"
二分图(能用 2 种颜色分开):细胞只是两两挨着 ,不会出现 3 个互相抱在一起。A--B--C--D 这种一条链、或者像棋盘一样的结构。规则: 只用 黑、白 2 种颜色 ,就能保证:相邻细胞颜色一定不一样
→ 这就是传统方法默认的 "理想情况"。
三角形(3 个细胞互相挨着) 是最常见的。
比如:A 挨着 B,B 挨着 C,C 又挨着 A → 三角环 ;这种结构2 着色绝对分不出来,必须用第 3 种颜色
##注释
基于这一真实复杂场景的观察,我们提出Disco (基于邻接感知协同着色的密集重叠细胞实例分割方法)------ 一种遵循分治 思想的邻接感知框架。它将数据驱动的拓扑标注策略 与约束式深度学习系统相结合,以解决复杂的邻接冲突。
- 显式标记策略:通过递归分解细胞图并分离出 "冲突集",将拓扑难题转化为可学习的分类任务。
- 隐式消歧机制:通过强制不同实例间的特征相异性,解决冲突区域的歧义,使模型学习到可分离的特征表示。
##注释
数据驱动的拓扑标注策略 + 显式标记:先靠数据(真实病理图)分析细胞的相邻关系,把细胞图里 2 色分不了的「三角环冲突细胞」单独揪出来,给它们贴个「冲突标签」,剩下的正常细胞用 2 色标注
递归分解细胞图 → 拆出冲突细胞 → 统一标为冲突集 → 转化为分类任务,AI 能直接学
约束式深度学习系统 + 隐式消歧:光标「冲突标签」还不够,这些冲突细胞还是挤在一起,AI 容易认错。所以给 AI 加一个强制约束 :只要是不同的细胞,哪怕都标了冲突,它们的特征也必须完全不一样 。让 AI 给每个细胞生成独一无二的特征,就算贴了一样的标签,也能精准区分开。
##注释
Disco 在GBC‑FS 2025 数据集上的PQ 指标显著提升 7.08% ,在所有数据集上平均提升2.72% 。此外,模型预测的冲突图 可作为一种全新的拓扑复杂度可解释工具,为数据驱动的病理研究提供新方向。代码已开源:https://github.com/SR0920/Disco。
引言
精准的细胞实例分割长期以来是细胞计数、形态学分析、空间组织学研究乃至癌症分级的核心任务 1。然而,受生物组织固有复杂性的影响,细胞常呈现出高度密集、严重重叠、形态多样 的特征,在保证高精度的前提下对每个独立细胞进行精准区分,是一项极具挑战性的工作。尽管基于目标检测 3、轮廓预测 4 与距离 / 方向映射 5 的主流方法已在特定场景取得显著进展,但这类方法均依赖局部像素或几何信息来判断像素所属的细胞实例。如图 1 (a-c) 所示,该思路在处理复杂细胞簇时会暴露固有的缺陷。这类方法的共性瓶颈在于缺乏显式建模细胞间全局拓扑约束的内在机制,其决策本质上是局部最优的,在复杂细胞簇中易产生系统性误差。
图一
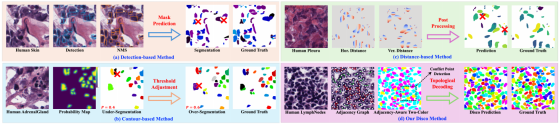
图 1 主流实例分割范式与本文提出的 Disco 框架的可视化对比
(a) 基于检测的方法受限于粗糙的边界框表示与启发式非极大值抑制(NMS)。
(b) 基于轮廓的方法对二值化阈值高度敏感。
(c) 基于距离的方法依赖复杂的后处理来完成实例重建。
(d) Disco 框架通过直接对细胞邻接图建模来重构问题,最终通过拓扑解码完成实例重建。
##注释
基于检测的方法:先框出每个细胞(Detection),再把框里的像素抠出来做分割(Mask Prediction),最后用 NMS 去掉重复的框。
弊端:
- 框不准:细胞挤在一起,框(Bounding Box)会盖住多个细胞。
- NMS 误杀 :为了去重,算法往往会把重叠部分的细胞预测直接删掉(图上红色叉叉),导致漏检。
结论 :靠框子和后处理,在细胞密密麻麻的地方必漏检。
基于轮廓的方法:先预测每个像素是细胞边缘还是内部(Probability Map),然后找阈值(Threshold)把边缘连起来,形成细胞轮廓。
弊端:
- 阈值极其敏感:
- 阈值设低一点(P=0.4):把背景噪音当细胞边缘,拆出一堆小碎片,造成过分割(Over-Segmentation)。
- 阈值设高一点(P=0.6):把细胞内部的缝隙漏掉,连不成完整轮廓,造成欠分割(Under-Segmentation)。
- 永远在 "过" 和 "欠" 之间摇摆。
结论 :靠单纯的像素边缘找细胞,在复杂组织里必拆分错误或粘连。
基于距离的方法:预测每个像素到细胞边界的水平距离(Hor. Distance)和垂直距离(Ver. Distance),或者预测中心距离,最后通过数学计算把这些距离 "拼回去" 成细胞实例。
弊端:
- 后处理极复杂:距离预测完了,还需要一套非常复杂的数学算法(Post Processing)才能把它们还原成细胞形状。
- 重叠处必乱套:两个细胞重叠时,它们的距离场会互相干扰,算法算不出来,导致最终预测混乱(红色叉叉处)。
结论 :靠距离场还原细胞,计算量大且在重叠区必乱套。
##注释
为突破上述局限,基于图着色理论的方法 6 近期应运而生,为该任务提供了具备全局拓扑感知能力的全新范式。其中,基于二分图理论的二着色方法 7 是最简洁高效的模型。与此同时,FCIS 方法 8 的开创性工作验证了通用四着色模型的可行性。但该范式的核心假设 ------ 二分图结构,在真实细胞拓扑中的有效性仍未得到验证,这是一个关键的研究空白。为严谨探究该问题,本文做出两项基础性贡献:第一,我们发布了GBC-FS 2025(2025 胆囊癌冰冻切片数据集) ,这是一个全新的大规模压力测试数据集,包含 2839 张胆囊癌冰冻切片图像,标注了 864204 个亚细胞级细胞核,呈现出高度复杂与密集的亚细胞结构。第二,基于该数据集与另外三个公开基准数据集,我们首次对细胞邻接图的着色特性开展系统性分析。如表 1 所示,我们的研究揭示了一个颠覆性发现:真实细胞图本质上为非二分图,普遍存在大量奇环(以三角环为主)。该发现表明,任何基于理想二分图假设的简易模型,在处理复杂病理图像时都存在根本性局限。这一结论亟需一种能够同时适配二分图与非二分图结构的全新方法。
表一
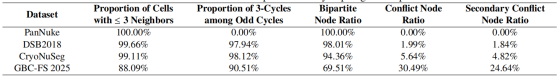
表1:关键拓扑属性的跨数据集比较
##注释
邻居数≤3 的细胞占比、奇环中 3 环(三角环)占比、二分图节点占比、冲突节点占比、二次冲突节点占比(「二次冲突节点」= 自己是冲突节点,且邻居也是冲突节点的细胞)
GBC-FS 2025:论文自建的高复杂度数据集
- 邻居数≤3 占比 88.09%:仅 88% 的细胞邻居数≤3,12% 的细胞有 4 个以上邻居,细胞极度密集
- 奇环中 3 环占比 90.51%:仍以三角环为主,但比例略降,出现少量更长的奇环,复杂度更高
- 二分图节点占比 69.51%:仅不到 70% 的细胞是二分图结构,超过 30% 的细胞是冲突节点
- 冲突节点 30.49%、二次冲突 24.64%:30% 的细胞是冲突节点,其中 24.64% 是二次冲突(冲突细胞扎堆)
- 结论:
- ,普遍存在大量奇环(以三角环为主)
##注释
针对这一严峻且被低估的挑战,我们提出Disco(基于邻接感知协同着色的密集重叠细胞实例分割方法) ,这是一个冲突感知的动态二着色框架。Disco 通过先进的图论着色思想,从底层重构了细胞实例分割的思路。我们未采用通用性更高的高着色数模型,因其会对占主体的简单二分图结构造成不必要的表征冗余。取而代之的是,我们提出更精细的分治 策略。Disco 框架的核心包含两项创新机制:显式标记 ,将拓扑冲突转化为可学习的分类目标;隐式消歧,在连续特征空间中解决离散标签的固有歧义。
##注释
隐式的巧办法 :完全不修改原来的离散标签(A、B、C 还是标「2」),只在连续特征空间里做区分,不用加新标签,靠特征本身消歧义,所以叫「隐式」
##注释
综上,本文的贡献可归纳为四点:
- 突破性拓扑分析:首次对真实细胞邻接图的着色特性开展系统性定量分析,颠覆性揭示其以密集 "冲突簇" 为特征的非二分图本质,为图着色范式应用于细胞分割提供了关键的数据驱动理论基础。
- 全新 Disco 框架:基于上述发现,提出遵循分治原则的冲突感知框架 Disco,创新性融合两大核心机制 ------ 显式标记将拓扑冲突转化为结构化可学习目标,隐式消歧通过端到端邻接约束在连续特征空间中解决离散标签歧义。
- 高密度案例研究数据集:发布 GBC-FS 2025 数据集,包含超 85 万个标注的亚细胞核实例。该数据集拥有前所未有的细胞密度与极端拓扑复杂度,冲突节点占比超 30%,为鲁棒分割算法的研发提供了不可或缺的压力测试平台。
- 最优性能与全新可解释范式 :Disco 在四个异构数据集上均取得当前最优(SOTA)性能,在 GBC-FS 2025 数据集上 PQ 指标显著提升 7.08%。此外,我们率先将预测得到的冲突图作为全新的拓扑量化工具,为数据驱动的病理研究开辟了新方向。
从局部线索到全局拓扑
细胞实例分割方法的发展历程,可被视作一种范式转变:从依赖局部几何信息的方法,转向能够建模全局拓扑结构的方法 9。本文将对现有方法展开综述分析,附录 A.2 则提供了更详尽的相关工作梳理。
局部线索的瓶颈
主流实例分割范式 11 尽管技术实现各异,但核心共性在于均依赖局部像素级或几何线索来推断实例归属。以 Mask R-CNN 12 为代表的基于检测的方法,依托粗糙的几何边界框进行运算,其最终性能往往受非极大值抑制(NMS)13 的启发式规则制约 ------ 在形态不规则、重叠度高的密集场景中,极易出现实例漏检问题(图 1 (a))。基于轮廓的方法 14 对二值化阈值高度敏感,阈值偏低会造成实例合并(欠分割),阈值偏高则会导致实例破碎(过分割)(图 1 (b))。StarDist 15、Hover-Net 16 等基于距离 / 方向的方法试图通过学习更丰富的特征表示缓解上述问题,却又依赖复杂且易出错的后处理算法从预测的向量场中重构实例,易引发误差传播,最终导致错误的实例分割(图 1 (c))。
这类方法的共性瓶颈在于缺乏内在机制来理解细胞间的全局拓扑结构。其决策本质上是局部最优的,在面对密集组织的全局复杂性时,不可避免地会出现系统性失效。
全局拓扑的潜力
为突破这一核心局限,学界涌现出一种全新思路:将实例分割问题抽象为图着色任务 17,以此显式建模全局拓扑约束。该范式中最简洁的实现形式是基于二分图理论的二着色18,理论上可处理所有无奇数长度环的图。这引发了一个关键且尚未验证的问题:真实场景中的细胞图在多大程度上符合这种理想的二分图结构?
与此同时,FCIS 8 的开创性工作验证了基于四色定理的通用四着色模型的潜力。但当底层拓扑结构较为简单时,该方法会引入不必要的特征表示冗余,还可能增加优化难度。这便构成了 "金发姑娘难题" 19:寻找一种复杂度恰到好处 的着色模型,既不过于简单,也不过于复杂。
##注释
金发姑娘走进熊家,尝粥、坐椅子、睡床,都要选不烫不凉、不大不小、不软不硬,刚刚好的那一个。
##注释
上述问题亟需通过系统性的拓扑分析,揭示细胞图的真实结构特性,这也成为本文探索最优、动态自适应着色方案的核心动机。
跨数据集拓扑分析
要提出严谨的解决方案,必须深刻理解问题的内在结构。尽管基于图的范式为实例分割提供了极具前景的新思路,但细胞邻接图的拓扑特性在很大程度上仍是未被探索的领域 20。为填补这一关键空白,我们在四个差异显著的数据集上开展了系统性的定量分析,以实证方式刻画真实生物组织的细胞拓扑特征。该分析不仅为本文提出的 Disco 框架奠定了坚实基础,也是本文的核心科学贡献之一。
图建模与拓扑预备知识
我们将细胞的空间排布形式化建模为细胞邻接图(Cell Adjacency Graph, CAG)21


该定理建立了图结构(奇环的存在性)与满足所有邻接约束所需最小颜色数之间的直接关联。如图 2 所示,细胞排布既可以形成简单的、可二着色的二分图结构,也可形成包含奇环的复杂非二分图结构。只要存在一个奇环,就至少需要三种颜色才能完成着色,这会导致所有二着色方案出现着色冲突 。在密集组织中,多个奇环会共享顶点,形成冲突簇 ;此时,多个引发冲突的节点之间也会相互邻接,我们将这种情况定义为二次冲突。接下来,我们将定量研究这些结构在真实数据中的分布情况。
图二
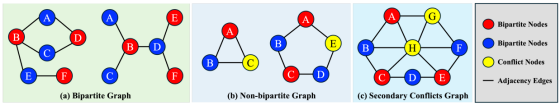
图 2 细胞邻接图的基本拓扑结构
(a) 可二着色的简单二分图结构:细胞排布比较简单,没有 3 个及以上细胞互相紧密挨在一起的情况,用两种「实例标签」(对应红 / 蓝)就能把所有相邻细胞完全分开
(b) 包含奇长度环(如三元环)的非二分图结构,该结构会引发着色冲突:3 个细胞互相紧密接触(形成 3 环 / 三角形),这时候用两种标签根本没法分配,总会有相邻细胞同标签,也就是论文里说的「着色冲突」
(c) 由相互连通的奇环构成的复杂 "冲突簇",会在相邻的冲突节点之间引发二次冲突:在极度密集的病理组织里,大量细胞互相紧密连接,形成了无数互相嵌套的奇环,冲突节点不是零散的,而是形成了密集的冲突簇
图特性的实证分析
我们分析了四个复杂度各异的数据集所对应的细胞邻接图,完整结果如图 3 所示。分析主要聚焦两个核心维度:局部连通性 与全局拓扑。
图三
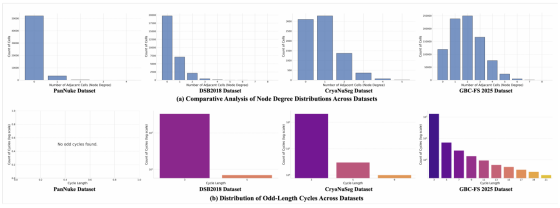
图 3 细胞邻接图拓扑结构的跨数据集对比分析
(a) 节点度分布:各数据集的局部连通性差异显著,从高度稀疏的 PanNuke 数据集,到呈密集簇状分布的 GBC-FS 2025 数据集。
(b) 奇长度环分布:三元环的广泛存在证实,绝大多数真实场景中的细胞图均为非二分图,其中 GBC-FS 2025 数据集的拓扑复杂度达到峰值。y 轴采用对数刻度。
局部连通性:节点度(即节点的邻接节点数量)23 反映了细胞的局部密集程度。图 3 (a) 展示了各数据集的节点度分布。尽管平均节点度较低表明图整体呈稀疏性,但长尾分布与最高达 8 的节点度,说明存在高度密集的局部邻域。如表 1 所示,在 DSB2018、CryoNuSeg 等简单数据集中,超过 99% 的细胞拥有不超过 3 个邻接节点;而在本文构建的更复杂的 GBC-FS 2025 数据集中,该比例降至 88.09%,证实局部连通性显著提升。
全局拓扑与非二分性:最具突破性的发现来自对奇环的分析。结果表明,除极度稀疏且完全为二分图的 PanNuke 数据集外,非二分图是真实细胞图的普遍特征。CryoNuSeg 数据集中 56.67% 的图像为非二分图,GBC-FS 2025 数据集中该比例为 29.17%。图 3 (b) 展示了这些奇环的长度分布,呈现出高度一致的规律:在所有非二分图数据集中,三元环(三角形)占全部奇环的比例超过 90%。这直接有力地证明,由三个及以上细胞直接接触形成的局部结构,是拓扑复杂度的主要来源 24。
着色冲突的普遍性
奇环的广泛存在 25 直接说明,简单的二着色模型无法适用。为量化这一问题,我们提出冲突节点的概念。

如表 1 的可行性分析所示,冲突节点比例在不同数据集间差异极大:PanNuke 数据集为 0%,而 GBC-FS 2025 数据集高达 30.49%。这意味着在复杂组织中,近三分之一的细胞存在拓扑冲突,无法通过二着色方案区分。此外,分析还揭示了更严峻的挑战 ------二次冲突。
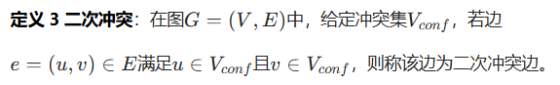
二次冲突分析表明,冲突节点并非孤立存在。所有包含非二分图的数据集均存在二次冲突,在极具挑战性的 GBC-FS 2025 数据集中,二次冲突节点比例达 24.64%。
这些定量结果得出明确结论:真实的细胞邻接图(尤其是密集病理图像对应的细胞图)本质上是非二分图,且以相互连接的奇环构成的密集冲突簇为典型特征。数据集分析的更多细节与可视化结果见附录 A.3。
方法
我们的拓扑分析揭示了真实场景中细胞邻接图(CAG)的核心本质特征:这类图既以简单的局部二分结构为主体,又穿插着关键的非二分冲突簇。为了更好地应对这种复杂多变的实际情况,我们提出了Disco ------ 一种全新的框架,其设计目标是在简单拓扑结构下保持高效,在复杂拓扑结构下保证鲁棒性。它摒弃了一体化的高色数解决方案,转而采用更精巧的「分而治之」思想,通过两大核心机制实现:用于生成标签的显式标记(Explicit Marking) ,以及用于模型优化的隐式消歧(Implicit Disambiguation)。我们的框架实现了完整的端到端学习流程,将拓扑分析、预测建模与约束学习协同融合,如图 4 所示。
图四
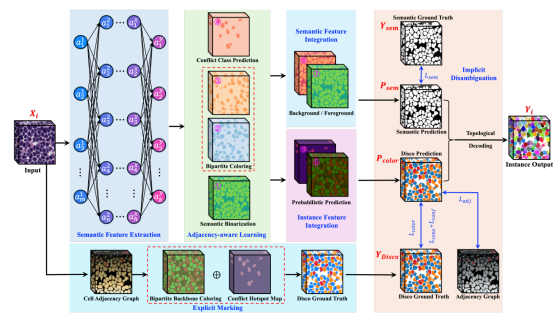
图 4 本文所提 Disco 方法的训练框架总览
该框架协同融合了三部分核心模块:
(1) 数据驱动的拓扑分析,生成 Disco 真值图YDisco,该真值图同时编码二分图结构与冲突热点区域;
(2) 双分支分割网络,学习预测基础语义图Psem与精细化的 Disco 着色图Pcolor;
(3) 解耦式损失体系,为模型提供针对性的监督信号。
至关重要的是,邻接约束损失(Ladj)依托真值邻接图,在连续概率空间中强制所有相邻实例的特征保持相异性,进而实现端到端的约束优化。
「分而治之」原则
Disco 方法的核心,是将「分而治之」策略应用于图着色问题。我们不再将细胞邻接图视为同质化的整体,而是首先根据其内在拓扑结构对问题空间进行分层。
从理论上看,图着色问题等价于将图G的顶点集V划分为最少数量的互不相交的独立集(Independent Set) {V1,V2,...,Vk}。尽管求解由色数χ(G)定义的这种最小划分是一个NP 难问题(NP-Hard Problem),但我们对数据集的实证分析得到了一个关键结论:细胞邻接图以一个规模庞大的二分子图为绝对主体 26。
##注释
图着色 = 把节点分成「最少的独立集」
一个独立集里的细胞,彼此都不挨着、不重叠 ,所以可以染同一种颜色 ;图着色的本质 :用最少的颜色 ,把所有节点分成几组独立集,每组一种颜色,保证相邻节点颜色不同。
色数χ(G):最少需要的颜色数
NP 难问题(NP-Hard):计算机算不出最优解;求「最少颜色数」的最优划分,是计算机无法快速求解的难题。无法在多项式时间内求出最优解
细胞邻接图以一个规模庞大的二分子图为绝对主体:真实病理图像里,90% 以上的细胞都属于二分主干 ,只有极少数细胞(复杂密集区)构成非二分的冲突区域。
##注释
这一关键结构特性,是我们「分而治之」原则的核心基础。它说明,一体化的高色数模型并非最优选择 ------ 它会将同样复杂的计算逻辑,同时应用于简单、庞大的二分区域,以及稀疏、复杂的非二分区域。相反,我们提出的最优策略是:首先高效处理占主导地位的二分分量,再针对孤立的、拓扑难度高的冲突节点,应用专门的处理机制。这一原则让我们能够设计出更具针对性、更高效的学习框架,具体细节将在后续章节展开。
显式标记:感知冲突的标签生成
「显式标记」策略将原始实例真值转换为融合拓扑信息的监督信号,该过程的可视化分解如图 5 所示。该策略通过一个高效的两阶段分解流程,将「分而治之」原则落地实现。
##注释
原始的实例分割真值:人工标注的细胞边界
融合了细胞拓扑结构信息的监督信号:用于模型训练的标注
两阶段分解流程:拓扑分解(拆分二分主干 + 冲突区域);标签生成(二分着色 + 冲突标记,生成 Disco 监督标签)
##注释
图五
图5. Disco标签生成过程的可视化分解示意图
我们的算法采用广度优先搜索(BFS),高效提取我们在分析中发现的最大二分子图。这一操作将绝大多数「简单」节点划分为两个大型独立集V1和V2,对应我们的两种基础颜色。

##注释
理论方法:对细胞邻接图无限递归做二分分解,用很多颜色给所有细胞着色,但这么做有两个致命问题:
- 递归分解的计算量爆炸,而且找「最小冲突集」本身就是NP 难问题
- 真实病理数据里,90% 以上的细胞都是能用 2 色解决的二分主干,只有极少数是拓扑冲突
基于充分表示原则,用 BFS(广度优先搜索)快速找出冲突细胞,直接给它们统一标第 3 种「冲突色」,不再递归分解。
Grem:去掉二分主干后,剩下的无法用 2 色区分的细胞组成的子图
冲突集* Vconf*:去掉二分主干后,剩下的所有拓扑冲突的细胞,合并成的一个集合
用基于 BFS(广度优先搜索)的高效启发式算法,来近似求解这个冲突集Vconf
BFS(广度优先搜索):经典的图遍历算法,能快速遍历图的节点,效率极高。
启发式算法 :不追求理论上的最优解,而是找一个足够好的近似解,速度极快,适合大规模数据。
最终生成的是(t+1)个取值的真值图YDisco:t=3,所以是 4 个取值:0(背景)、1(二分色 1,橙色)、2(二分色 2,蓝色)、3(冲突色,红色)
##注释
隐式消歧:解耦式约束损失体系
尽管「显式标记」策略提供了强有力的监督信号,但它在「二次冲突」区域存在理论上的歧义性局限。为解决这一问题,我们提出了「隐式消歧」机制,通过全新的端到端损失体系,在连续特征空间中弥补离散标签的不足。
##注释
二次冲突区 = 两个冲突细胞(都标红色)还互相挨着
歧义:标签一样(都是红色)→ 但实际是两个细胞 → 离散标签(固定 0/1/2/3)没法区分
隐式消歧:不靠固定标签,而是在连续特征空间 (模型学到的小数特征向量),用损失函数强制:只要相邻,特征就必须不一样。就算标签都是红色,特征也完全不同 → 模型能分清是两个细胞
之前给细胞标 "橙 / 蓝 / 红" 三色标签的方法(显式标记)虽然好用,但在两个冲突细胞挨在一起 的地方,会出现标签分不清的 bug。为修好这个 bug,我们做了 "隐式消歧",用新的训练规则,在模型的特征数据里强行让相邻细胞不一样,解决标签分不清的问题。
##注释

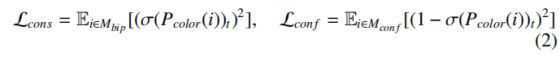
##注释
一致性损失 Lcons:
Mbip 是正常二分区域,绝对不应该被标成冲突色(红色),因此我们希望 σ(Pcolor(i)t)(像素 i 是冲突类的概率)尽可能接近 0。
损失项是「概率的平方」:概率越接近 0,损失越小;概率越大,惩罚越重。
冲突消解损失 Lconf:
Mconf 是冲突区域,必须被标成冲突色(红色),因此我们希望 σ(Pcolor(i)t) 尽可能接近 1。
损失项是「(1 - 概率) 的平方」:概率越接近 1,损失越小;概率越小,惩罚越重。
两个损失一推一拉,完美配合:
- Lcons 防止「正常区错标红色」,避免把简单问题复杂化;
- Lconf 保证「冲突区必标红色」,避免漏标拓扑复杂区域;
##注释

从概念上看,Ladj可被理解为一种监督对比损失:对于每个实例,同一实例内的其他像素构成正样本集,分类损失Lcolor会推动这些像素的特征凝聚;反之,图中该实例的所有相邻实例,均被视为显式负样本。通过最小化与这些负样本的余弦相似度,Ladj推动模型学习到这样的特征流形:不同实例(尤其是相邻实例)的表征在角度上被最大程度地分离。
这实现了强大的双重功能:
- 在二分区域,它推动模型学习显式、正交的二色编码;
- 在二次冲突区域,它为着色损失提供反向梯度,迫使模型在次级特征维度中学习可分离的表征。
该机制的有效性在我们的特征空间可视化中得到了充分验证,如图 6 所示。各损失分量的详细公式见附录 A.5。
图六
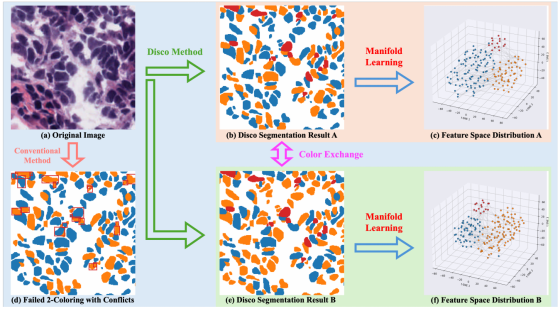
图6. 隐式消歧可视化与着色歧义鲁棒性分析。通过t-SNE算法将每个细胞实例的最终4D概率向量投影至三维空间,直观呈现Disco框架的核心"隐式消歧"机制及其对着色非唯一性的不变性特征。
##注释
t-SNE:经典流形学习降维算法,用于将高维特征投影到低维(3D)空间,直观展示特征的聚类分布;
4D 概率向量:每个细胞实例对应 4 个概率值,分别对应「背景、二分色 1、二分色 2、冲突色」4 类,是模型输出的实例特征;
三簇特征完全分离、无重叠,蓝 / 橙簇正交(角度完全分开),完美验证了「隐式消歧」的效果:
- 邻接约束损失Ladj强制相邻细胞特征正交,所以不同类、相邻的细胞特征在空间里完全分开,不会混淆;
- 推拉损失Lcons+Lconf保证了正常区 / 冲突区的正确分类,特征聚类清晰。
着色非唯一性的不变性特征:Disco 模型学到的是细胞实例的本质特征,而不是人为的颜色标签。哪怕二分色的标签交换,特征的拓扑结构完全不变,证明了模型对着色非唯一性的强鲁棒性。
##注释
实验
数据集
为全面评估 Disco 方法的性能与泛化能力,我们在4 个公开及自研的高异质性数据集上开展实验。这些数据集涵盖多样的成像模态、细胞类型与广泛的拓扑复杂度区间。所有数据集在进入处理流程前,均预处理为无重叠的 256×256 像素块。实验所用数据集具体为:PanNuke 28、DSB2018 29、CryoNuSeg 30 与 GBC-FS 2025。
PanNuke 包含 7901 张 256×256 图像,源自 19 种人体器官的 H&E 染色、福尔马林固定石蜡包埋(FFPE)组织切片,覆盖 5 种不同细胞类型,总计标注 189744 个实例。我们遵循其官方划分策略,使用折 1 与折 2 用于训练与验证,折 3 用于最终测试。
DSB2018 包含 670 张尺寸不一(256×256 至 520×696)的荧光显微图像,采用 DAPI 与 Hoechst 染色,总计标注 29443 个实例。我们将数据集划分为 536 张训练图、67 张验证图与 67 张测试图。
CryoNuSeg 包含 30 张来自 10 种人体器官、尺寸为 512×512 的 H&E 染色冷冻切片图像,总计标注 8178 个实例。我们将图像裁剪为 256×256 块,随后划分为 96 个训练块、12 个验证块与 12 个测试块。
GBC-FS 2025 为本研究首次提出,专为高密度案例研究设计,用于对算法鲁棒性进行压力测试。该数据集源自一张经深度标注的胆囊癌冷冻切片全视野数字切片(WSI),对复杂拓扑的处理能力提出了前所未有的挑战。数据集包含 2839 张 256×256 的 H&E 染色图像,标注 864204 个亚细胞核对实例。数据集划分为 2271 个训练块、284 个验证块与 284 个测试块。更多细节见附录 A.6。
实现细节与评估指标
所有实验基于 PyTorch 框架开展,在搭载 8 块 NVIDIA RTX 4090 GPU 的服务器上运行。优化器采用 Adam,初始学习率设为 1×10⁻⁴,权重衰减为 5×10⁻⁴。采用阶梯式学习率衰减策略,在第 70 轮训练时将学习率缩小 10 倍;前 100 次迭代使用线性预热,保证训练初期稳定。所有模型均训练 200 轮。
分割性能采用以下指标评估:戴斯系数(Dice) 31、聚合杰卡德指数(AJI) 32、检测质量(DQ) 33、分割质量(SQ)* 与* 全景质量(PQ)。本文所有表格中,最优性能分数以红色粗体标注,次优分数以蓝色粗体标注。
评估与结果
定量与定性评估
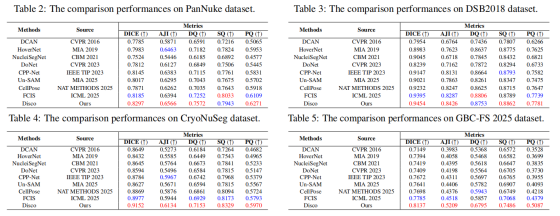
为验证 Disco 方法的有效性与泛化能力,我们在 4 个拓扑复杂度各异的异质数据集上,与多种主流实例分割方法开展全面的定量与定性对比。对比方法包括:基于检测的 DoNet 3;基于轮廓预测的 DCAN 4 与 NucleiSegNet 34;基于距离映射的 HoverNet 16、CPP-Net 35 与 CellPose 36;基于 SAM 的基础模型 Un-SAM 37;以及基于图论的 FCIS 8。如表 2--5 所示,Disco 方法在全部 4 个数据集上均展现出一致且优异的性能。
表二~五
定量对比
在拓扑最简单、细胞邻接图 100% 为二分图的 PanNuke 数据集上,本方法在关键指标 AJI 与 PQ 上取得最优结果,分别为 65.66% 与 62.71%。这证明即便在最简单场景下,本动态着色框架可自动退化为高效的 2 着色模型,性能优于需学习更复杂表征的通用 4 着色模型 FCIS。
随着拓扑复杂度提升,Disco 的优势愈发显著。在中等复杂度的 DSB2018 与 CryoNuSeg 数据集上,Disco 在 AJI 与 PQ 指标上均全面领先。值得注意的是,在 DSB2018 上,尽管 FCIS 在 DQ 指标上略有优势,但本方法在 SQ 上显著更优,表明本模型生成的细胞轮廓更精准,最终在综合指标上占优。
该优势在极具挑战性的 GBC-FS 2025 数据集上达到顶峰。该数据集充斥密集的冲突簇 与二次冲突 ,对所有方法均构成严苛考验。表 5 结果清晰显示,Disco 在该数据集上性能碾压式领先。本方法 AJI 达到 52.09%,相较次优方法 FCIS 绝对提升 6.91%、相对提升 15.3%。同时,Disco 在 DQ 与 SQ 维度均全面领先。这无可辩驳地证明,本研究的显式标记 + 隐式消歧策略在处理真实场景中极端拓扑复杂度时,具备无与伦比的鲁棒性与有效性。
可视化对比
我们在图 7 中给出分割结果的直观可视化对比。这些可视化结果展示了 Disco 在染色方式、细胞形态与密度差异显著的 4 个数据集上精准分割细胞的能力。在白色框标注的高难度区域,Disco 相较次优方法 FCIS 展现出明显优势,可成功分离紧密簇集的实例。更多可视化结果见附录 A.7。
图七
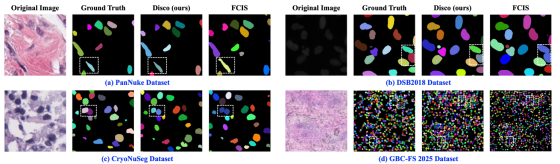
图7. 四个数据集分割结果的定性比较。各子图从左至右依次显示:输入图像、真实标注结果、本研究Disco方法的预测结果以及FCIS方法的预测结果。
消融实验
为系统验证 Disco 方法各核心设计的有效性,我们在难度最高的 GBC-FS 2025 数据集上开展了全面的消融实验。
框架级对比
我们首先进行框架级对比,以验证本动态着色方案的优越性,结果如表 6 所示。结果明确表明,无冲突解决机制的纯 2 着色基线性能最差(AJI 为 37.85%),进一步印证了前文关于处理非二分结构必要性的结论。相较于 FCIS 的通用 4 着色方案,Disco 框架在 PQ 指标上实现 7.08% 的显著绝对提升。我们认为该优势源于本研究的分治策略,它为网络提供更清晰、更具针对性的学习目标,从而避免通用高色数模型中潜在的优化困难与表征冗余。
表六
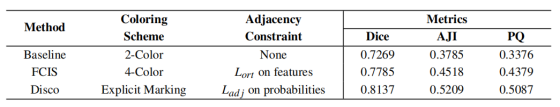
表6: GBC -FS 2025不同着色框架消融研究
损失分量分析
我们进一步拆解 Disco 的损失函数体系,验证各分量的贡献,结果如表 7 所示。实验结果表明,本文设计的所有损失项均对最终性能有正向贡献。其中,邻接约束损失(L_adj)起到决定性作用;仅在基础显式标记模型中加入该损失,即可使 PQ 绝对提升 6%(从 42.57% 升至 48.26%)。该结果从定量角度有力证明,本文提出的、通过隐式消歧解决二次冲突的 L_adj,是整个框架成功的核心驱动力。当所有损失分量协同作用时,最终 Disco 模型在 PQ 上达到 50.87% 的最优性能,表明本文设计的各组件均具备必要性与有效性。
表七
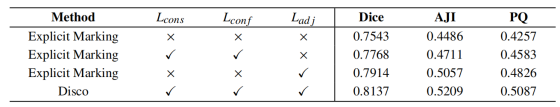
表7:基于 GBC -FS 2025平台的Disco损失系统组件消融研究
结论
通过首次对细胞邻接图的着色特性开展系统分析,本文揭示了真实细胞邻接图具有显著的非二分性 ,阐明了简单 2 着色模型在该领域的理论局限。为解决该挑战,本文提出 Disco 框架,一种基于分治原则的全新架构。
Disco 采用显式标记 策略,动态且智能地将细胞图分解为庞大的二分主干与稀疏的冲突集。更重要的是,它通过具备端到端邻接约束的损失体系实现隐式消歧 。该机制通过在连续特征空间中学习可分离表征,解决由二次冲突引发的离散标签歧义。
大量实验表明,Disco 在多个异质数据集上达到当前最优性能,并具备出色的泛化能力。此外,本文开创性地将预测的冲突图作为新型可解释性工具,为拓扑复杂度量化提供新路径。综上,Disco 为复杂细胞实例分割问题提供了理论严谨、高效且可解释的全新范式。
相关工作
基于检测的实例分割
以 Faster R-CNN 13 为代表的开创性工作提出了 "先检测后分割" 范式,并由 Mask R-CNN 12 拓展至实例分割任务。该两阶段方法先为单个实例生成边界框 38 候选,再在定位区域内完成分割。其固有的实例分离能力使其成为广泛使用的框架,尤其适用于半监督细胞分割任务 39。然而,该范式的性能从根本上受限于对轴对齐边界框的依赖,这类框对形态复杂或不规则的细胞表征过于粗糙。此外,最终实例集合往往由非极大值抑制(NMS)的启发式规则决定,在密集、重叠的细胞簇中易造成有效检测被错误抑制,这一关键失效模式如图 1 (a) 所示。
基于轮廓与距离映射的分割
为规避边界框的局限,第二类主流方法致力于学习编码实例身份的稠密逐像素表征。
基于轮廓的方法旨在显式预测细胞间边界。经典的 U-Net 40 通过在损失函数中对细胞边缘加权来辅助边界学习,后续再通过分水岭等后处理方法勾勒实例。该架构对领域产生了深远影响。后续工作聚焦于提升边界预测精度,例如 DCAN 4 引入专用边界类别通道。UNet++41 的嵌套跳跃连接、FullNet 42 与 CIA-Net 43 的多尺度上下文聚合等架构优化进一步改善了边界勾勒效果。尽管取得这些进展,基于轮廓的方法仍对预测边界图的二值化阈值高度敏感,导致在实例合并与碎裂间难以权衡,如图 1 (b) 所示。
基于距离映射的方法通过学习更丰富的空间关系实现更鲁棒的分离。StarDist 15 与 CellPose 36 分别学习预测从像素到细胞边界或中心的向量。Hover-Net 16 拓展了这一思路,同时预测水平与垂直距离图。这类方法虽达到当前最优性能,但通常依赖复杂且易出错的后处理 44 来重建实例,易引发误差传播 45------ 预测向量场中的微小偏差会被放大为严重的实例分割错误,如图 1 (c) 所示。尽管从 CPP-Net 35 到基于视觉 Transformer 的 CellViT 46 等近期模型持续推动该范式发展,它们仍普遍依赖局部几何线索解码。
分割中的图论方法
上述所有范式共有的理论瓶颈是:缺乏内在机制对细胞间的全局拓扑约束进行建模。为解决该问题,一种将任务抽象为图着色问题的全新视角应运而生。
图着色是图论中的基础问题,与调度、资源分配问题联系紧密。图 G=(V,E) 的 k 着色是一个映射 C:V→{1,...,k},满足对任意边 (u,v)∈E 均有 C (u)≠C (v)。满足该条件的最小 k 值称为图的色数 χ(G)。
计算机视觉早期工作将图着色 47 作为区域合并的后处理步骤,而将其融入端到端学习则是近期的创新。受四色定理 48(任意平面图满足 χ(G)≤4)启发,FCIS 8 首次提出训练网络直接预测细胞图的 4 着色。该开创性工作证明了将实例分割重新建模为受拓扑规则约束的多分类问题的巨大潜力。
比四色定理更基础的概念是 2 着色,根据柯尼希定理,2 着色对任意二分图均成立。这引出了一个关键的 "金发姑娘问题":真实细胞拓扑所需的最小且足够的颜色数是多少 49?简洁优雅的 2 着色模型是否足够,还是需要 4 着色模型的通用鲁棒性?
本文工作致力于回答这一核心问题,并据此提出最优解决方案。我们不将细胞图视为套用现有理论的被动对象,而是将其视作独特且未被探索的结构。首先,通过系统拓扑分析,我们首次从实验上回答该问题,揭示 "近二分但含密集冲突簇" 的复杂现实。其次,基于该发现,我们设计一种全新的非迭代机制,通过图感知损失系统将强拓扑先验注入深度网络。因此,本文工作不仅解决了 "金发姑娘问题",还为如何将深刻的图论原理与深度表征学习结合以解决结构化预测任务提供了全新视角。
补充分析与可视化
为更全面、直观地阐释本文核心动机与所提着色策略的有效性,我们在图 A.1 中对全复杂度跨度的数据集进行了并列可视化对比。该图可视化分解了从原始输入到最终着色结果的完整分析流程,为全文核心论点提供直接证据。
图A.1
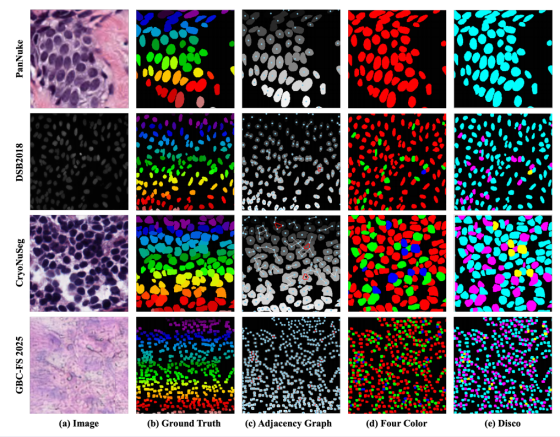
图A.1. 细胞图拓扑结构与着色策略的跨数据集综合可视化展示。每行对应一个特定数据集,拓扑复杂度从上至下递增。
列 (a) 为输入图像,列 (b) 为实例真值。这两列确立了问题背景,展现了不同基准数据集在成像模态(H&E、荧光)、细胞密度与形态上的显著异质性。(b) 中随机着色的实例图代表理想分割目标。
列 (c) 为带奇环检测的细胞邻接图(CAG)。该列可视化了拓扑分析的核心:将实例图抽象为底层图结构,节点为细胞质心,边代表邻接关系。我们对每张图进行关键诊断:属于非二分组件的边标为红色,简单二分组件的边保留灰色。该可视化直观揭示了 "拓扑复杂度谱系":PanNuke 数据集的图全为灰色,证实其为完美二分图;DSB2018 与 CryoNuSeg 开始出现红色边,表明存在孤立或小簇奇环;而 GBC-FS 2025 数据集则呈现大规模连通的红色冲突边网络,直观体现密集 "冲突簇" 与 "二次冲突" 的普遍性。
列 (d) 为通用贪心 4 着色。该列展示了通用贪心着色算法的结果。一个关键实验发现是:在全部四个数据集(包括高度复杂的 GBC-FS 2025)中,三种颜色在实验上足以解决所有邻接约束。该证据强烈表明,真实细胞图的实际色数通常为 χ(G)=3。因此,任何基于四色定理、为最坏情况(色数为 4)设计的框架都会引入不必要的表征冗余与潜在优化困难。
列 (e) 为 Disco 着色方案。该列展示了所提 Disco 框架的自适应性与靶向高效性。在完美二分的 PanNuke 数据集上,算法自动退化为最优 2 着色方案,避免颜色浪费;在中等复杂度的 DSB2018 与 CryoNuSeg 数据集上,Disco 有效采用冲突感知 2 着色策略,用两种主色(蓝、紫)高效着色庞大的二分主干,用专用冲突色(黄)精准稀疏标记 (c) 中识别的少量拓扑冲突;在极端复杂的 GBC-FS 2025 上,Disco 稳健执行相同的结构化冲突感知着色方案。这为模型提供了清晰、带拓扑信息的监督信号,显式定位大量冲突热点,为 "隐式消歧" 机制在特征空间中解决剩余歧义奠定基础。
综上,该全面可视化有力地并列验证了本文核心论点:Disco 并非单纯的着色方法,而是为真实细胞图像拓扑特性量身定制的、原理化的数据驱动框架。它既避免了纯 2 着色的理论不足,也规避了 4 着色的实际冗余,在效率、鲁棒性与可解释性之间取得最优平衡。
细胞图色数的理论与实验边界
4.2 节已说明我们终止图分解的务实选择。本节对细胞邻接图(CAG)的色数 χ(G) 展开更深入的理论与实验讨论,进一步支撑本文设计理念。
色数上界的理论说明
图论中一个经典结论给出了基于图最大度 Δ_G 的色数上界。

实验发现:"3 色现象"
最引人注目的实验发现(如定性结果图 A.1 所示)是:在全部四个异质数据集(包括拓扑复杂的 GBC-FS 2025)中,三种颜色始终足以解决所有邻接约束。这表明,真实细胞图若为非二分图,其实际色数几乎总是 χ(G)=3;若为二分图则 χ(G)=2。这一 "3 色现象" 与理论上界 9 形成鲜明对比。
细胞图低色数结构起源的假说
低色数的一致观察引出更深层问题:细胞图为何在结构上如此受限?我们提出假说:这并非巧合,而是支配组织构成的基本物理与生物学原理的结果。
图论表明,高色数(χ(G)≥4)要求图包含 K₄子式 ------ 即可收缩为 4 顶点完全图 K₄的子图。K₄子图对应四个不同细胞实例两两相互邻接的情况,如图 A.2 所示。
图 A.2
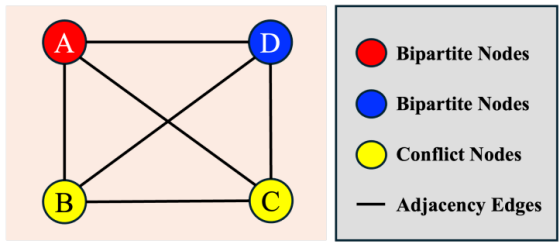
图A.2. 四顶点完全图(K4)示意图及其在细胞分割中的生物物理解释。K4子图代表一种拓扑结构配置,其中四个独立细胞实例(A、B、C、D)彼此完全相邻。我们推测,在准二维组织切片中形成此类结构在生物物理层面极不可能,这源于细胞体积和膜张力的物理限制。现实世界细胞图中缺乏类似K4子图这类高阶冲突结构,为经验观察到的"三色现象"提供了理论基础。
我们认为,该结构在准二维组织切片中生物物理上不可行。受细胞体积、膜张力与最优填充原则(类似于圆填充问题)限制,四个不同的近似凸状物体很难实现两两同时物理接触。尽管三向接触(形成 3 环 / 三角形)在细胞连接中常见,但四向相互接触是拓扑不稳定且能量不利的状态。
Disco 的结论与依据
该假说为实验发现的 "3 色现象" 提供理论支撑。它表明,细胞排列的内在生物物理特性天然阻止了需要 4 种及以上颜色的高阶冲突结构(如 K₄子式)形成。该发现是 Disco 框架的最终依据:无需为生物物理上不现实的高色数最坏情况(如 4 着色方案)做准备,本文 "2+1" 方案完美适配观测现实 ------ 用两种颜色高效处理占主导的二分结构,用一种足够的冲突色解决常见的低阶(以 3 环为主)拓扑冲突。
损失系统的详细公式
Disco 框架的端到端优化由解耦约束损失系统驱动。如公式 (1) 所述,总损失为五个分量的加权和。本节给出各分量的详细公式与设计依据。
符号说明:设 P_sem∈ℝ^(B×2×H×W) 为语义对数几率,P_color∈ℝ^(B×(t+1)×H×W) 为网络输出的着色对数几率,对应批次大小为 B 的图像。设 Y_sem∈{0,1}^(B×H×W) 为二元语义真值,Y_Disco∈{0,...,t}^(B×H×W) 为 Disco 真值图,本文中 t=3。设 σ(・) 为通道级 Softmax 函数,Ω 为图像所有像素坐标集合。
基础损失 L_sem 与 L_color
这两个损失为网络双分支提供核心监督信号。
语义损失 L_sem监督基础前景 / 背景分割,由逐像素交叉熵损失 L_CE 与区域级戴斯损失 L_Dice 线性组合而成:
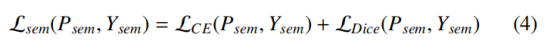
该损失确保网络精准定位所有细胞区域的空间范围。
着色损失 L_color监督多分类 Disco 标注任务,由加权交叉熵 L_WCE 与戴斯损失组成:
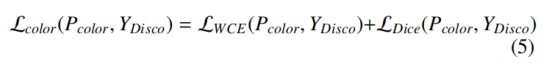
像素 i 的加权交叉熵定义为:
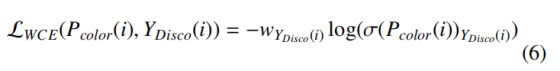
其中 w_c 为类别 c 的权重。本文将冲突类权重 w_t 显著高于二分颜色权重(w_t≫w_c,c∈{1,...,t−1}),强制模型优先学习这些关键拓扑特征。
解耦正则损失 L_cons 与 L_conf
为落实 "分治" 方案的特定逻辑,我们引入一对互补正则损失,构成 "推拉" 机制。
二分一致性损失 L_cons为 "拉" 力,惩罚在拓扑简单的二分区域 M_bip={i∈Ω | Y_Disco (i)∈{1,...,t−1}} 内错误预测冲突色的情况,目标是保证冲突类的稀疏性与靶向使用:
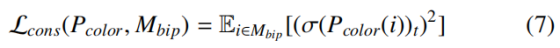
冲突消解损失 L_conf为 "推" 力,奖励在拓扑冲突区域 M_conf={i∈Ω | Y_Disco (i)=t} 内高置信度预测冲突类,确保关键信号不被抑制:
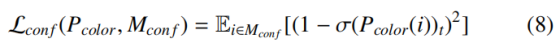
用于隐式消歧的邻接约束损失 L_adj
这是 "隐式消歧" 机制的核心,旨在解决二次冲突区域中离散标签的理论局限。它作用于网络输出的连续特征流形,将不适定的离散问题转化为适定的特征分离问题。
设 s_k 为实例 k 的像素坐标集合。首先定义该实例的平均概率向量 P̄(s_k)∈ℝ^(t+1),为其所有像素 Softmax 概率向量的均值:
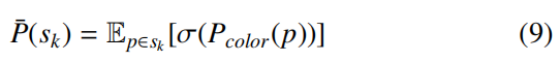
该向量代表实例在概率空间中的平均特征嵌入。
邻接约束损失 L_adj定义为细胞邻接图 G 上所有边 E 的全局势能函数,通过最小化相邻实例特征表征的余弦相似度,最大化其角度分离度:
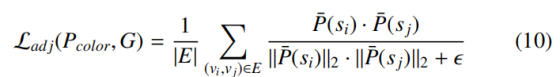
其中‖・‖₂为向量 L2 范数(欧几里得范数),ϵ 为数值稳定小常数 10⁻⁸。
该损失为核心分类目标提供强反向梯度。在二次冲突区域,L_color 驱使相邻实例归为同一离散类别 t,而 L_adj 则通过强制网络在次级通道(c∈{1,...,t−1})学习不同激活模式,将二者的连续表征分离。从概念上讲,这可视为一种监督对比损失,图邻接关系定义了需在特征空间中相互排斥的负样本对。该机制确保即便离散标签存在歧义,实例在学习到的特征流形上仍可分离,这对拓扑复杂区域的精准实例重建至关重要。
GBC-FS 2025 高密度案例研究数据集
本节全面介绍本文首次构建并提出的 GBC-FS 2025(胆囊癌冷冻切片 2025)数据集,详细阐述其临床动机、数据构建流程、关键特征与领域潜在价值。
高密度病理分割的关键空白
胆囊癌(GBC)是最常见的胆道恶性肿瘤,占比 80%--95%,位列主要消化系统癌症之一。因其起病隐匿、缺乏特异性早期症状,多数患者确诊时已至晚期,预后极差:转移性患者 5 年生存率低于 5%,中位生存期常不足 6 个月。这一现状凸显了对高质量细胞与亚细胞级定量分析的迫切需求,以推动早期病理特征识别、免疫浸润分析与空间异质性表征的突破。
然而,胆囊癌的组织学特征极具挑战:肿瘤细胞常形成边界模糊的致密巢状结构,免疫反应则在肿瘤周边诱发密集淋巴细胞簇与三级淋巴结构(TLS),形成密度极高、重叠严重的区域。在这些区域,传统细胞分割方法频繁失效,导致下游分析偏差,如免疫浸润评估失真、生物标志物发现不可靠。
现有公开数据集(如 MoNuSeg、PanNuke、Lizard/CoNIC)虽有价值,但多聚焦常见癌症,缺乏亚核粒度标注。因此,主流模型在胆囊癌这类高密度复杂组织上泛化性差。GBC-FS 2025 数据集专为填补该空白构建,提供首个大规模、胆囊癌冷冻切片亚核标注基准。
数据集构建与亚细胞标注
(1) 数据来源:GBC-FS 2025 的 2839 张 256×256 图像源自一位 71 岁 IVB 期腺癌患者的胆囊癌冷冻切片全视野数字切片(WSI)。冷冻切片天然存在染色不均、冰晶伪影、组织变形与核膜模糊 / 不完整等问题,无法可靠完成细胞级核标注。因此,我们不标注难以稳定识别的完整细胞,转而标注视觉上稳定可重复的染色质样致密亚细胞聚集体。其余患者的完整 WSI 数据将在后续发布。
(2) 标注方法:所有实例由四名接受基础形态学训练的标注人员使用 QuPath(v0.5.1)软件手动勾勒,并由病理学背景博士审核。对高度模糊区域,咨询病理学家进行有限样本验证,确保整体标注方向符合冷冻切片基本形态特征。此外,标注目标为 "染色质样致密亚细胞结构",这类结构在含噪冷冻切片中比完整细胞核更易稳定识别。该选择旨在构建反映该数据模态真实视觉挑战的基准,而非模拟更清晰的 FFPE 标注。
(3) 质量控制:鉴于冷冻切片边界天然模糊,我们采用多轮专家审核流程,比传统 IoU / 戴斯指标更适配该场景。标注由初级人员完成后,经病理学背景博士审核;不确定区域标记后由团队资深成员裁决,少数疑难案例提交病理学家抽样检查。
(4) 组织构成:病理学家指导的区域评估显示,数据集约含 89.69% 癌细胞区域、6.11% 成纤维细胞区域与 4.20% 免疫细胞区域。
该数据集在三方面具有实用意义:生物学上,助力探索分裂核与核碎裂,量化异质性与多极性,为肿瘤侵袭性与预后评估提供依据;算法上,亚核标注提供比传统细胞级标签更细粒度的监督信号,提升在密集、重叠、形态复杂区域的鲁棒性,是分割算法压力测试的理想数据集;临床上,亚核勾勒既可作为核分割基准,也可辅助细胞分割预处理 ------ 核单元聚合可近似全细胞边界,支撑免疫浸润分析、肿瘤分级与预后评估。
关键特征与现有数据集对比
GBC-FS 2025 区别于现有数据集的核心不仅在于聚焦罕见且具挑战的癌症类型,更在于其前所未有的拓扑复杂度与亚核标注粒度。表 8 直接对比了本文所用其他数据集,以明确其独特性。
表八
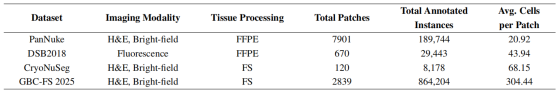
表8:实验所用数据集概述。 FFPE 表示福尔马林固定石蜡包埋样本,FS表示冰冻切片样本。
表格清晰显示,GBC-FS 2025 处于 "拓扑复杂度谱系" 的极端位置:平均实例密度远超其他数据集,每块 304.44 个实例,是次密数据集 CryoNuSeg 的四倍以上。该极高密度是其复杂图结构的主要成因(正文第 3 节分析)。其冲突节点比例高达 30.49%,二次冲突节点比例达 24.64%,远超其他公开基准一个数量级。
超大规模(86 万 + 实例)与极端拓扑复杂度的独特结合,使 GBC-FS 2025 成为领域不可或缺的资源。它作为现有数据集的关键补充,为最具挑战的高密度分割场景提供急需基准,支撑 Disco 这类鲁棒拓扑感知算法的开发与严格压力测试。
补充定性结果
为更全面直观地证明 Disco 框架的鲁棒性与泛化能力,我们在图 A.3 中展示四个数据集上的大量定性分割结果。该图展示了 Disco 在各类挑战场景下的性能,验证其在不同成像模态、染色方式、细胞形态与密度下的一致精度。
图A.3
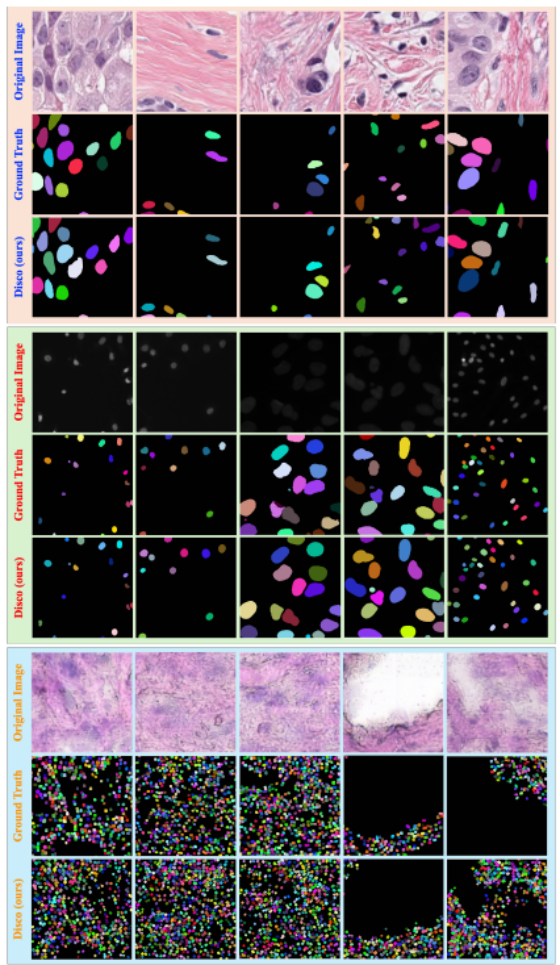
图A.3展示了Disco方法在四个数据集上的定性分割结果对比图。每个数据集区块中,首行呈现原始输入图像,中间行显示对应的真实实例掩码,末行展示Disco方法的最终分割预测结果。为便于观察,所有实例掩码均采用随机颜色进行可视化呈现。
这些可视化结果证明 Disco 能出色适应差异极大的视觉域,在明场 H&E 染色图像与荧光显微图像上表现相当;同时成功处理全密度区间的细胞,从稀疏排列到极端密集拥挤环境。在高难度区域,Disco consistently 生成与真值高度一致的实例掩码,精准勾勒不规则形状,分离紧密簇集的细胞。
一个尤其值得注意的发现是,在标注不完全的案例中,仅用现有标签训练的 Disco 展现出强大泛化能力:成功识别并分割大量真值中完全缺失的亚细胞核。对比对应面板中的 Disco 行与真值行可清晰观察到,本方法正确勾勒出大量额外有效实例。这种 "补全" 能力凸显了学习到的拓扑先验的鲁棒性,彰显了 Disco 在半监督 / 弱监督学习场景下的潜力,显著提升其在穷尽标注难以实现的真实应用中的实用价值。
计算效率对比
分析证实,本文方法的图相关组件(CAG 构建、BFS 标注)为一次性离线预处理步骤,推理时无额外开销。因此推理速度仅由神经网络架构与后处理决定。
为公平对比推理效率,我们在单张 NVIDIA RTX 4090 GPU 上对所有模型进行基准测试,输入为 256×256。结果汇总于表 9。
表九
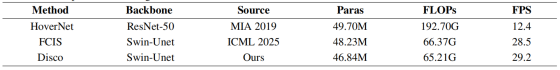
表9:计算成本比较。Params表示模型参数。FLOPs为256×256输入尺寸下的运算量。FPS表示推理过程中的每秒帧数。
结果清晰证明 Disco 框架的高效性:推理速度达 29.2 FPS,不仅极具竞争力,还略快于图论强基线 FCIS(28.5 FPS);同时比 HoverNet 等依赖高计算量后处理的方法快两倍以上。
值得注意的是,即便采用复杂的双分支头与解耦损失系统,Disco 模型参数量仍低于 FCIS(46.84M vs 48.23M),计算量更低(65.21G vs 66.37G FLOPs)。这些结果无可辩驳地表明,Disco 达到当前最优精度并非依靠大幅增加计算复杂度,而是通过更高效、更靶向的学习策略。本方法在不牺牲、甚至部分提升实际计算效率的前提下,成功实现性能提升。
L_adj 与 NonAdjLoss 的对比
NonAdjLoss 51 与本文 L_adj 在概念上相似 ------ 均利用预定义邻接图惩罚网络输出中不期望的关系,但二者的目标、机制与应用域存在根本差异,这也凸显了本文方法的独特性。
(1) 目标根本不同:NonAdjLoss 的目标是语义级正确性,通过阻止不同语义类别间不可能的邻接关系来保证解剖一致性,是提升区域纯度与语义正确性的损失。与之相对,本文 L_adj 的目标是实例级可区分性,强制同一语义类别内不同实例的特征相异。前者提升区域纯度,后者提升实例可分离性。
(2) 运行机制不同:NonAdjLoss 作用于像素级,惩罚局部邻域内禁止类别概率的同时出现;而本文 L_adj 是作用于实例级的监督对比损失,先将每个实例抽象为平均特征向量,再将邻接图中的边视为 "负样本对",直接最小化这些对的特征相似度。这种基于对比学习、在实例级强制特征流形分离的机制,是 NonAdjLoss 不具备的。
(3) 应用域与解决问题不同:NonAdjLoss 解决 "拓扑错误的语义布局" 问题;本文 L_adj 解决 "密集簇中实例身份歧义" 问题,尤其是基于图着色的实例分割特有的 "二次冲突" 挑战。
综上,尽管两种方法均使用邻接图,却用于解决完全不同维度的问题,采用本质不同的机制:NonAdjLoss 确保不同类别在不该接触时不接触,而本文 L_adj 确保同一类的不同实例在接触时特征可分离。
不同着色策略的公平对比
本文 "2+1" 方案看似引入中间歧义,实则比始终使用三种颜色的静态贪心 3 着色更优、更高效、更具 "动态自适应性"。依据有二:
(1) 动态自适应且更高效的标注策略:标准贪心 3 着色无论输入图简单或复杂,始终尝试使用三种颜色;与之相对,本文 "显式标记" 算法动态自适应 ------ 面对简单二分细胞图(如 PanNuke 数据集)时,自动退化为最优 2 着色方案,仅用两种颜色,完全避免引入第三种;仅在检测到不可避免的拓扑冲突时,才 "激活" 并引入专用冲突色。这种 "按需分配" 策略不仅理论上更优雅,还为网络提供更简洁的监督信号,符合奥卡姆剃刀原则,避免简单场景下的表征冗余。
(2) 驱动更鲁棒的特征学习:"标签歧义" 是刻意的设计选择,结合本文 L_adj,可产生建设性张力,驱动更鲁棒特征流形的学习。干净的 3 着色监督仅要求网络模仿离散标签,而本文 "2+1" 方案迫使模型利用次级特征维度解决着色损失与邻接损失间的冲突,从而学习真正可分离、具备拓扑感知的特征表征。
为实验验证该核心论点,我们开展相关消融实验:在 GBC-FS 2025 数据集上对比 Disco、标准贪心 3 着色与 FCIS 方法。表 10 结果显示,采用 "显式标记 + 隐式消歧" 策略的 Disco 框架显著优于在干净无歧义 3 着色与 4 着色标签上训练的模型。这一大幅性能差距提供有力实验证据,证明本文看似 "迂回" 的设计实则是更有效的学习策略,引导模型获得更鲁棒、更精准的解。
表十
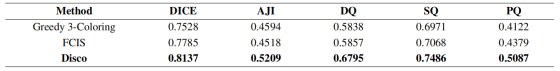
表10: GBC -FS 2025不同着色策略的公平性比较