Structural, functional and evolutionary diversity of 4-coumarate-CoA ligase in plants
植物中4-香豆酸辅酶 A 连接酶(4CL)的结构、功能与进化多样性
CYP450综述-20年-地表最强系列-文献精读-4_cyp76ah-CSDN博客
紫杉醇的合成生物学技术路线综述--文献精读195_a synthetic biology roadmap for sustainable produc-CSDN博客
4-香豆酸:CoA连接酶晶体-文献精读138_苯丙素生物合成通路4cl-CSDN博客
摘要
4 - 香豆酸辅酶 A 连接酶(4CL)参与调控不同苯丙烷类生物合成途径的代谢通量。4CL 的表达在植物发育阶段受到精细调控,并对生物胁迫、非生物胁迫等环境信号作出响应。该酶在姜黄素、白藜芦醇、生物燃料合成及营养品质改良等代谢途径工程中具有重要应用价值。为实现基于 4CL 的目标代谢物高效规模化生产,必须对其功能与表达水平的调控机制进行系统深入研究。
苯丙烷途径为植物中大量次生代谢产物合成提供前体。在该途径中,4 - 香豆酸辅酶 A 连接酶(EC 6.2.1.12,4CL)是生成活化硫酯的关键分支点酶。作为通用苯丙烷途径中三个共有步骤的最后一个酶,它负责为不同苯丙烷类物质分流前体。在植物中,4CL 以多种同工酶形式存在,由一个小的基因家族编码。该酶属于腺苷酸形成酶家族,催化将羟基或甲氧基取代的肉桂酸衍生物转化为相应硫酯的反应。这些硫酯进一步用于苯丙烷类化合物的生物合成,而苯丙烷类物质因其广泛的营养与药用价值备受关注。此外,研究者已在多种植物中对 4CL 进行了功能鉴定,揭示其在植物生长发育及应对生物、非生物胁迫中的作用。同时,特定 4CL 同工酶在不同外界刺激下呈现差异表达调控,进而使代谢通量转向特定代谢物的合成。进化研究表明,4CL 在单子叶植物与双子叶植物分化后发生了独立进化。本文对 4CL 进行了全面综述,涵盖其进化、功能、基因 / 蛋白结构、在代谢物合成与细胞区域化中的作用及调控机制。基于现有研究,我们探讨了利用 4CL 开展代谢途径工程的应用前景。
引言
植物体内存在种类繁多的代谢产物,这些物质对植物正常的生长、发育和繁殖至关重要。代谢产物在植物生命周期中发挥着关键作用,包括机械支撑、吸引传粉者与食果动物、抵御生物和非生物胁迫、与环境互作以及化感作用等。植物次生代谢产物可依据化学结构、组成、溶解性及其合成途径进行分类。例如,按化学结构可分为三大类:(1)萜类化合物;(2)苯丙烷类化合物;(3)生物碱。近年来代谢物分析技术的快速发展,使得人们能够在细胞与细胞器水平上大规模研究代谢产物、单一代谢物的衍生物及其代谢通路。已有大量次生代谢产物被鉴定,这些物质具有着色、增香、药用等诸多重要特性与应用价值。
苯丙烷类化合物由苯丙氨酸、酪氨酸等芳香族氨基酸经苯丙烷途径 合成。除上述芳香族前体氨基酸外,色氨酸也在植物质体中经莽草酸途径 合成。莽草酸途径利用糖酵解产生的磷酸烯醇式丙酮酸与己糖单磷酸途径产生的 4 - 磷酸赤藓糖合成芳香族氨基酸。经典苯丙烷途径起始于苯丙氨酸解氨酶(PAL)对苯丙氨酸、酪氨酸解氨酶(TAL)对酪氨酸的氧化脱氨,分别生成肉桂酸和对香豆酸。在后续步骤中,肉桂酸在羟化酶与甲基转移酶的催化下,转化为对香豆酸、咖啡酸、阿魏酸、5 - 羟基阿魏酸、芥子酸等多种甲氧基与羟基衍生物。这些衍生物经 4CL 催化生成对应的辅酶 A 酯。4CL 的催化产物进一步在各类氧化酶、还原酶与转移酶的作用下,参与木质素、类黄酮、花青素、橙酮、芪类、香豆素、木栓质、角质、孢粉素等物质的合成。因此,4CL 是苯丙烷途径中的关键分支点酶之一 。编码该酶的基因家族包含多个 4CL 同工型,这些同工型可能行使多样的功能,但其具体作用仍有待全面解析。目前已发表的苯丙烷途径相关综述中,关于 4CL 的系统介绍较少。本文对 4CL 的现有研究进行全面综述,并对其进化规律及作为代谢工程靶标酶的未来应用前景进行展望。
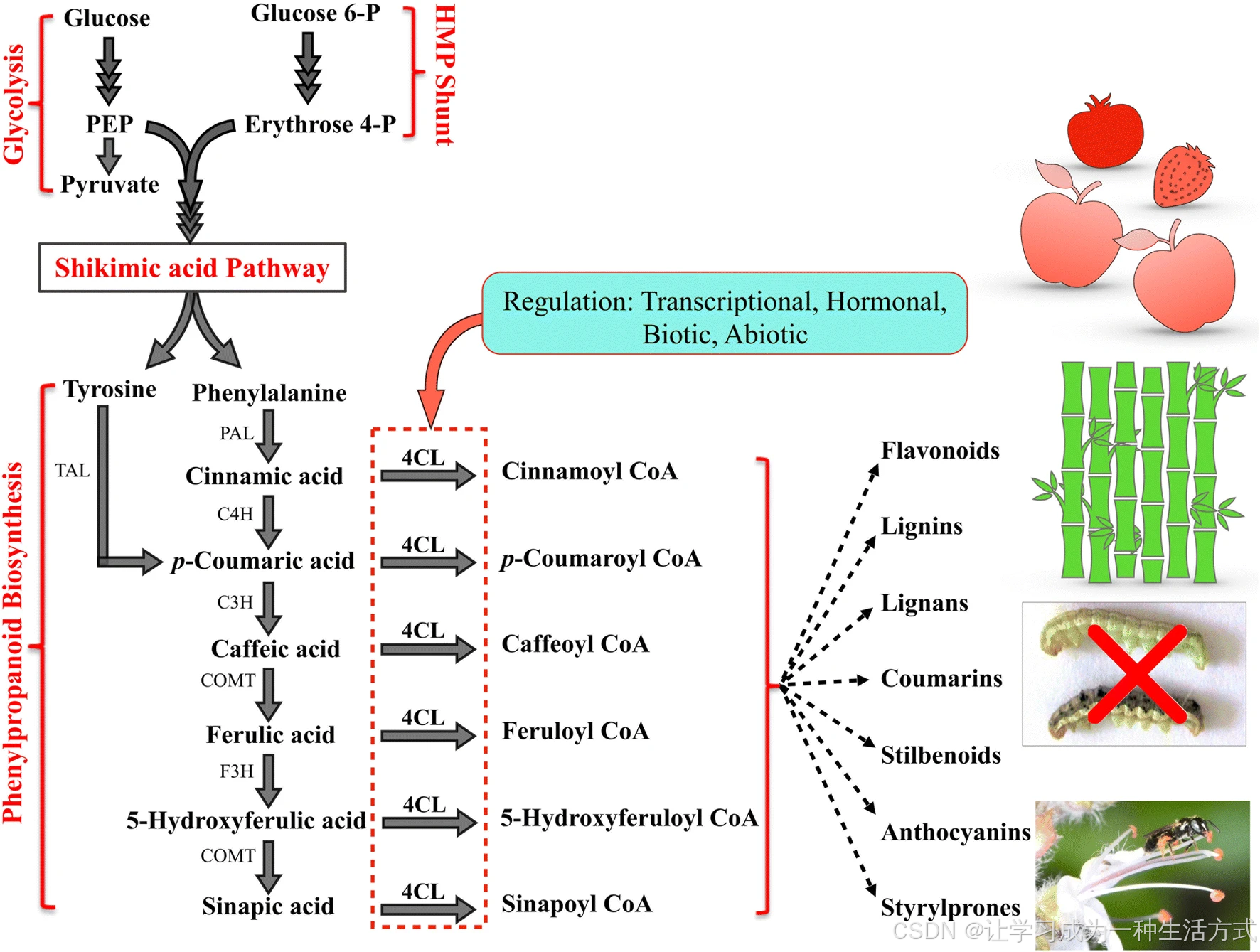
苯丙烷通用途径及 4CL 作为分支点关键酶的作用 HMP shunt:己糖单磷酸旁路4CL:4 - 香豆酸辅酶 A 连接酶PEP:磷酸烯醇式丙酮酸PAL:苯丙氨酸解氨酶TAL:酪氨酸解氨酶C4H:肉桂酸 4 - 羟化酶C3H:对香豆酸 3 - 羟化酶COMT:咖啡酸 O - 甲基转移酶F5H:阿魏酸 5 - 羟化酶
植物 4CL 酶独特的催化特性
4CL 催化辅酶 A(CoA)与肉桂酸及其甲氧基 / 羟基衍生物(如咖啡酸、阿魏酸)的连接反应(图 1)。该酶属于腺苷酸形成酶家族 ,含有两个保守肽基序:框 I(SSGTTGLPKGV)和框 II(GEICIRG) 。框 I 为腺苷一磷酸(AMP)结合结构域,在腺苷酸形成酶家族所有成员中均高度保守。该结构域广泛存在于所有生物的蛋白质中,参与多种功能,例如果蝇荧光素酶、短杆菌肽 S 合成酶、各类 CoA 连接酶等。而框 II 的功能尚不明确,且不直接参与催化反应。
此外,** 底物结合结构域(SBD)** 在不同 4CL 同工酶中具有高度变异性。在拟南芥中,研究者采用结构域交换法研究了两个同工酶 At4CL1 和 At4CL2 的底物特异性决定区域。两种酶均能以对香豆酸为底物,但只有 At4CL1 可利用阿魏酸。研究鉴定出两个相邻的结构域,分别称为 SBD I 和 SBD II;其中任意一个来自 At4CL1 的 SBD 均足以识别阿魏酸。At4CL2 的底物结合口袋(SBP)包含 12 个氨基酸残基,该结果基于短杆菌肽 S 合成酶的同源模型并通过晶体结构解析确定。这些氨基酸包括:Ile-252、Tyr-253、Asn-256、Met-293、Lys-320、Gly-322、Ala-323、Gly-346、Gly-348、Pro-354、Val-355 和 Leu-356。这 12 个氨基酸位于保守的框 I 和框 II 基序两侧。
酰基活化酶超家族成员间序列一致性较低,但均含有保守的框 I。在拟南芥中,该超家族包含 77 个 4CL 家族基因;系统发育分析显示它们分为 7 个类群,且均具有独特的保守框 I。
酶的潜在功能可通过一级序列与三维结构分析进行解析。研究者利用定点突变、结构域交换、分子建模、晶体学等手段解析了 4CL 的多种结构特征。毛白杨 Pt4CL1 的晶体结构以萤火虫荧光素酶为模型,通过反常散射法与分子置换法解析获得。Pt4CL1 包含 536 个氨基酸,由两个球状结构域组成:N 端结构域(434 个氨基酸)和 C 端结构域(102 个氨基酸)。N 端结构域又分为 N1、N2、N3 三个子结构域;N1 与 N2 结构相似,中心均含有 6 个平行 β- 折叠与 2 个反平行 β- 折叠,8 个中心 β- 折叠的两侧分别排布 4 个和 2 个 α- 螺旋。3 个残基对 Pt4CL1 催化活性至关重要(Lys-438、Gln-443、Lys-523),另有 5 个残基参与底物结合(Tyr-236、Gly-306、Gly-331、Pro-337、Val-338)。
基于晶体结构及其他植物中可转化芥子酸的 4CL 报道,研究者通过结构域交换与定点突变对 At4CL2 进行改造,以提升其对芥子酸的转化效率。改造分为两步:(1)用 At3g21230 基因编码的 4CL 样蛋白的底物结合口袋,替换 At4CL2 中从框 I 到框 II 的底物结合口袋区域;(2)单独缺失 V355 或 L356(不同时缺失)。研究者还替换了 At4CL2 中的 3 个氨基酸残基(N256A、M293P、K320L);改造后的 4CL 对芥子酸的催化效率提升了 30 倍。
部分 4CL 可通过蛋白间互作实现调控功能。例如,毛果杨的 Ptr4CL3 与 Ptr4CL5 同工酶可相互作用,形成由 3 个 Ptr4CL3 和 1 个 Ptr4CL5 组成的四聚体。研究者构建数学模型分析不同比例下的动力学特征与激活组分,并结合显微切割、免疫共沉淀、化学交联、双分子荧光互补与质谱等实验分析,证实 Pt4CL5 在 Ptr4CL3-Ptr4CL5 四聚体复合物中发挥调控作用。
烟草 Nt4CL2 同工酶的催化机制也得到了研究。Nt4CL2 的晶体结构显示,该酶在催化过程中呈现两种构象:腺苷酸形成构象 与硫酯形成构象 。催化过程中,酶先后通过两种构象将底物转化为腺苷酸中间体,再进一步生成硫酯形式。
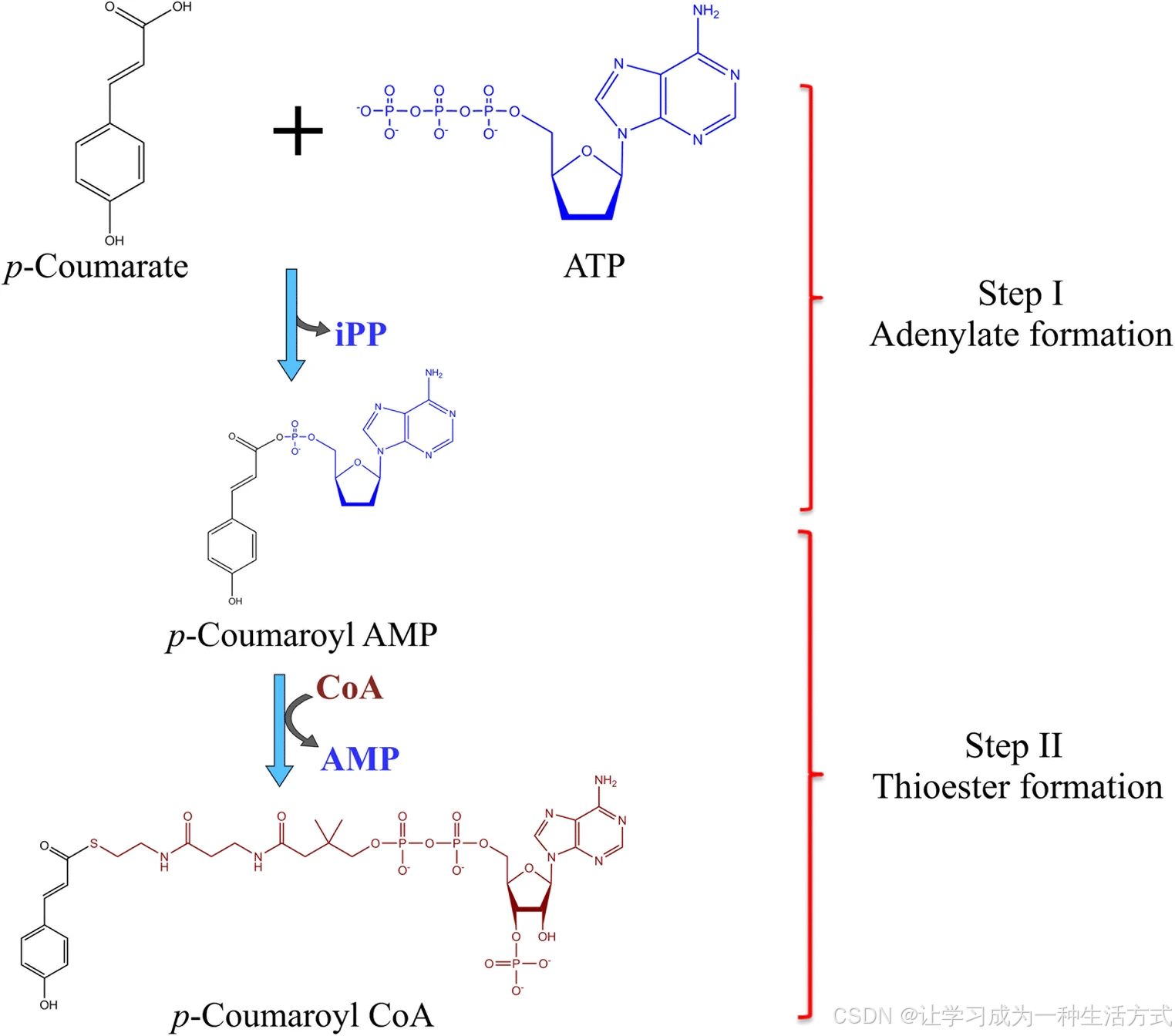
4CL 催化的反应机制 4CL 通过两步反应将对香豆酸转化为对香豆酰辅酶 A:第一步,形成腺苷酸中间体;第二步,进而形成硫酯产物。
植物 4CL 的进化与多样性
苯丙烷途径的进化是陆生植物成功登陆定植的关键前提。该途径的产物如类黄酮可抵御紫外线,木质素则为植物提供结构支撑。对 4CL 基因的系统发育分析表明,4CL 基因在单子叶植物与双子叶植物中分化为两个独立的进化枝。这可能是由于在进化过程中,单 / 双子叶植物分化后,4CL 基因发生了独立进化。
在双子叶植物中,4CL 基因分为两大类:I 型与 II 型。I 型主要参与单木质醇(木质素前体)生物合成 ;II 型则参与除木质素以外的其他苯丙烷类物质合成。在拟南芥中,At4CL1、At4CL2 和 At4CL4 属于 I 型,At4CL3 属于 II 型。
有研究将单子叶植物的 4CL 划分为另外两类:III 型与 IV 型,功能分别与双子叶 I 型、II 型类似。在水稻中,Os4CL1、Os4CL3、Os4CL4、Os4CL5 属于 III 型,Os4CL2 属于 IV 型。这些不同类群的形成表明,4CL 基因在双子叶与单子叶植物中的分化是各自独立发生的。
然而,裸子植物火炬松(Pinus taeda )的 Pinta4CL3,在进化上更接近被子植物 II 型 4CL,而非同属裸子植物的 III 型 Pinta4CL1。这提示 I 型与 II 型的分化可能早于裸子植物与被子植物的分化。
拟南芥 At4CL1 与 At4CL2 的氨基酸序列相似度更高(一致性 86%),而 At4CL3 与二者的一致性仅分别为 71% 和 73%,说明 At4CL1 和 At4CL2 是较晚进化出的基因。类似地,丹参基因组中有 10 个 Sm4CL 相关基因,系统发育分析显示只有 3 个(Sm4CL1、Sm4CL2、Sm4CL3)属于真正的 4CL 家族:Sm4CL1 与 Sm4CL2 属于 I 型,参与木质素合成;Sm4CL3 属于 II 型,参与类黄酮合成。
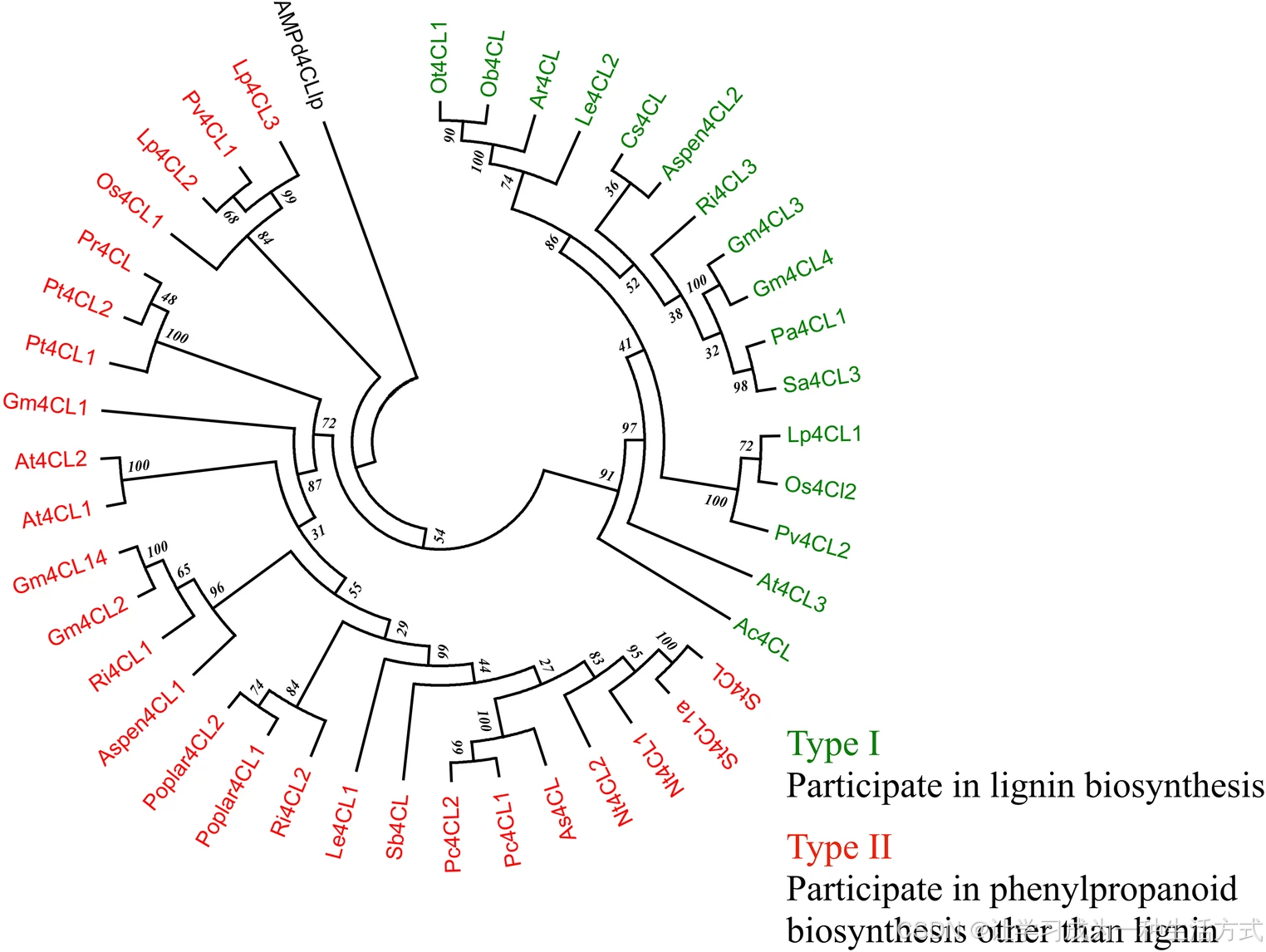
显示不同 4CL 蛋白进化关系的系统发育树
I 型类群参与木质素合成,II 型类群参与木质素以外的苯丙烷类物质生物合成。
该系统发育树基于推导氨基酸序列,采用 ** 邻接法(NJ)** 构建。仅选取已完成功能鉴定的 4CL 蛋白序列用于建树。
(序列比对:Muscle;进化模型:p-distance;自展检验:1000 次重复)
4CL:4 - 香豆酸辅酶 A 连接酶
用于构建系统发育树的 4CL 氨基酸序列登录号及物种名称:
拟南芥 Arabidopsis thaliana AMPd4CLlp(AMP 依赖型 4CL 样蛋白):NM_123172
拟南芥 A. thaliana At4CL1:U18675
拟南芥 A. thaliana At4CL2:AF106086
拟南芥 A. thaliana At4CL3:AF106088
藿香 Agastache rugosa Ar4CL:AY587891
洋葱 Allium cepa Ac4CL:AY541033
当归 Angelica sinensis As4CL:AMP18194.1
茶树 Camellia sinensis Cs4CL:DQ194356
大豆 Glycine max Gm4CL1:AF279267
大豆 G. max Gm4CL2:AF002259
大豆 G. max Gm4CL3:AF002258
大豆 G. max Gm4CL4:X69955
大豆 G. max Gm4CL14:X69954
紫草 Lithospermum erythrorhizon Le4CL1:D49366
紫草 L. erythrorhizon Le4CL2:D49367
黑麦草 Lolium perenne Lp4CL1:AF052221
黑麦草 L. perenne Lp4CL2:AF052222
黑麦草 L. perenne Lp4CL3:AF052223
烟草 Nicotiana tabacum Nt4CL1:U50845
烟草 N. tabacum Nt4CL2:U50846
水稻 Oryza sativa Os4CL1:X52623
水稻 O. sativa Os4CL2:L43362
圣罗勒 Ocimum sanctum Ot4CL:HM990148
罗勒 Ocimum basilicum Ob4CL:KC576841
柳枝稷 Panicum virgatum Pv4CL1:EU491511.1
柳枝稷 P. virgatum Pv4CL2:JF414903
欧芹 Petroselinum crispum Pc4CL1:X13324
欧芹 P. crispum Pc4CL2:X13325
火炬松 Pinus taeda Pt4CL1:U12012
火炬松 P. taeda Pt4CL2:U12013
辐射松 Pinus radiata Pr4CL:ACF35279.1
杂交杨 Populus hybrida Poplar4CL1:AF008184
杂交杨 P. hybrida Poplar4CL2:AF008183
颤杨 Populus tremuloides Aspen4CL1:AF041049
颤杨 P. tremuloides Aspen4CL2:AF041050
甜樱桃 Prunus avium Pa4CL1:GU990523
覆盆子 Rubus idaeus Ri4CL1:AF239687
覆盆子 R. idaeus Ri4CL2:AF239686
覆盆子 R. idaeus Ri4CL3:AF239685
黄芩 Scutellaria baicalensis Sb4CL:BAD90937.1
花楸 Sorbus aucuparia Sa4CL3:GU949553
马铃薯 Solanum tuberosum St4CL1:M62755
马铃薯 S. tuberosum St4CL1a:AF150686
|---------------------------|------------------------------------|----------------------------------------|-----------------|------------------|--------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------|
| Plant | Annotated 4CLs | Characterized 4CLs | Class-I | Class-II | References |
| Arabidopsis thanliana | 14 | 4 | 3 | 1 | The Arabidopsis Genome Initiative ([2000](#Plant Annotated 4CLs Characterized 4CLs Class-I Class-II References Arabidopsis thanliana 14 4 3 1 The Arabidopsis Genome Initiative (2000) and Costa et al. (2005) Salvia miltiorrhiza 10 3 2 1 Wang et al. (2015) Populus trichocarpa 20 6 5 1 Zhang et al. (2015) Populus pruinosa 20 5 4 1 Zhang et al. (2015) Populus euphratica 20 5 4 1 Zhang et al. (2015) Salix suchowensis 12 4 3 1 Zhang et al. (2015))) and Costa et al. ([2005](#Plant Annotated 4CLs Characterized 4CLs Class-I Class-II References Arabidopsis thanliana 14 4 3 1 The Arabidopsis Genome Initiative (2000) and Costa et al. (2005) Salvia miltiorrhiza 10 3 2 1 Wang et al. (2015) Populus trichocarpa 20 6 5 1 Zhang et al. (2015) Populus pruinosa 20 5 4 1 Zhang et al. (2015) Populus euphratica 20 5 4 1 Zhang et al. (2015) Salix suchowensis 12 4 3 1 Zhang et al. (2015))) |
| Salvia miltiorrhiza | 10 | 3 | 2 | 1 | Wang et al. ([2015](#Plant Annotated 4CLs Characterized 4CLs Class-I Class-II References Arabidopsis thanliana 14 4 3 1 The Arabidopsis Genome Initiative (2000) and Costa et al. (2005) Salvia miltiorrhiza 10 3 2 1 Wang et al. (2015) Populus trichocarpa 20 6 5 1 Zhang et al. (2015) Populus pruinosa 20 5 4 1 Zhang et al. (2015) Populus euphratica 20 5 4 1 Zhang et al. (2015) Salix suchowensis 12 4 3 1 Zhang et al. (2015))) |
| Populus trichocarpa | 20 | 6 | 5 | 1 | Zhang et al. ([2015](#Plant Annotated 4CLs Characterized 4CLs Class-I Class-II References Arabidopsis thanliana 14 4 3 1 The Arabidopsis Genome Initiative (2000) and Costa et al. (2005) Salvia miltiorrhiza 10 3 2 1 Wang et al. (2015) Populus trichocarpa 20 6 5 1 Zhang et al. (2015) Populus pruinosa 20 5 4 1 Zhang et al. (2015) Populus euphratica 20 5 4 1 Zhang et al. (2015) Salix suchowensis 12 4 3 1 Zhang et al. (2015))) |
| Populus pruinosa | 20 | 5 | 4 | 1 | Zhang et al. ([2015](#Plant Annotated 4CLs Characterized 4CLs Class-I Class-II References Arabidopsis thanliana 14 4 3 1 The Arabidopsis Genome Initiative (2000) and Costa et al. (2005) Salvia miltiorrhiza 10 3 2 1 Wang et al. (2015) Populus trichocarpa 20 6 5 1 Zhang et al. (2015) Populus pruinosa 20 5 4 1 Zhang et al. (2015) Populus euphratica 20 5 4 1 Zhang et al. (2015) Salix suchowensis 12 4 3 1 Zhang et al. (2015))) |
| Populus euphratica | 20 | 5 | 4 | 1 | Zhang et al. ([2015](#Plant Annotated 4CLs Characterized 4CLs Class-I Class-II References Arabidopsis thanliana 14 4 3 1 The Arabidopsis Genome Initiative (2000) and Costa et al. (2005) Salvia miltiorrhiza 10 3 2 1 Wang et al. (2015) Populus trichocarpa 20 6 5 1 Zhang et al. (2015) Populus pruinosa 20 5 4 1 Zhang et al. (2015) Populus euphratica 20 5 4 1 Zhang et al. (2015) Salix suchowensis 12 4 3 1 Zhang et al. (2015))) |
| Salix suchowensis | 12 | 4 | 3 | 1 | Zhang et al. ([2015](#Plant Annotated 4CLs Characterized 4CLs Class-I Class-II References Arabidopsis thanliana 14 4 3 1 The Arabidopsis Genome Initiative (2000) and Costa et al. (2005) Salvia miltiorrhiza 10 3 2 1 Wang et al. (2015) Populus trichocarpa 20 6 5 1 Zhang et al. (2015) Populus pruinosa 20 5 4 1 Zhang et al. (2015) Populus euphratica 20 5 4 1 Zhang et al. (2015) Salix suchowensis 12 4 3 1 Zhang et al. (2015))) |
研究者对松科落叶松属(Larix )11 个物种的 4CL 基因进化进行了分析,比较了其基因拷贝数、GC 含量、密码子使用偏好、序列分化程度并开展系统发育分析。结果显示,4CL 序列可分为两个旁系同源进化枝:4clA 和 4clB 。这两个分支均包含所研究的 11 种落叶松的序列,但部分物种在同一分支中存在多条序列。上述结果表明,4clA 与 4clB 这两个姐妹分支起源于落叶松共同祖先的基因复制事件 ,从而使两个等位基因在全部 11 个物种中共同存在。基因复制发生后,二者可能发生了独立进化,表现为同义替换、非同义替换、核苷酸替换的平均距离以及转换 / 颠换比值均存在差异。由此可见,4CL 基因家族在物种分化过程中发生了基因复制与缺失,这可能是等位基因序列与数量存在差异的潜在原因。据此推测,松科植物中可能存在2~3 个主要的 4CL 基因位点 。
本文更新了多个植物物种中已注释并经功能鉴定的 4CL 基因列表,包括拟南芥、丹参、毛果杨、灰杨、胡杨和簸箕柳等(表 1)。在上述物种中,尽管注释的 4CL 基因多达 10~20 个,但仅有 3~6 个完成了功能验证。
植物 4CL 的基因结构与基因组定位
已有大量植物的 4CL 基因被研究,结果表明它们多以小型基因家族 形式存在,编码序列完全相同、高度相似或分化明显的蛋白。在拟南芥中,通过全基因组生物信息学分析共注释出 14 个推定的 4CL 基因,其中仅 11 个开展了功能鉴定;其余 3 个基因含有过氧化物酶体靶向序列(基于 C 端信号肽分析),而典型的 4CL 基因不具备这一特征。
拟南芥 4CL 基因在染色体上分布广泛:
- 1 号染色体:At4CL1、At4CL3、At4CL9、At4CL10
- 3 号染色体:At4CL2、At4CL5、At4CL8
- 4 号染色体:At4CL6、At4CL7
- 5 号染色体:At4CL4、At4CL11
其中只有 At4CL2 和 At4CL5 在 3 号染色体上成簇分布。在这些基因中,At4CL1、At4CL2、At4CL3 和 At4CL5 具有功能活性 。
序列分析显示:
- At4CL1 和 At4CL2 含有3 个内含子 ;
- At4CL3 含有6 个内含子 。At4CL3 多出的 3 个内含子中,第一个由打断第一个外显子产生,另外两个则由打断 At4CL1/At4CL2 的第二个外显子产生。
At4CL1、At4CL2、At4CL3 的 5′非翻译区与推定启动子序列存在差异。三者翻译起始密码子 ATG 上游分别在−124、−103、−116 bp 位置存在推定的 TATA 框。
对于 PAL、C4H、4CL 等苯丙烷途径相关基因,其启动子区通常含有保守的 P 元件、A 元件和 L 元件。At4CL1 和 At4CL2 的启动子仅含 P 盒和 L 盒,而 At4CL3 同时含有全部三个保守元件 。
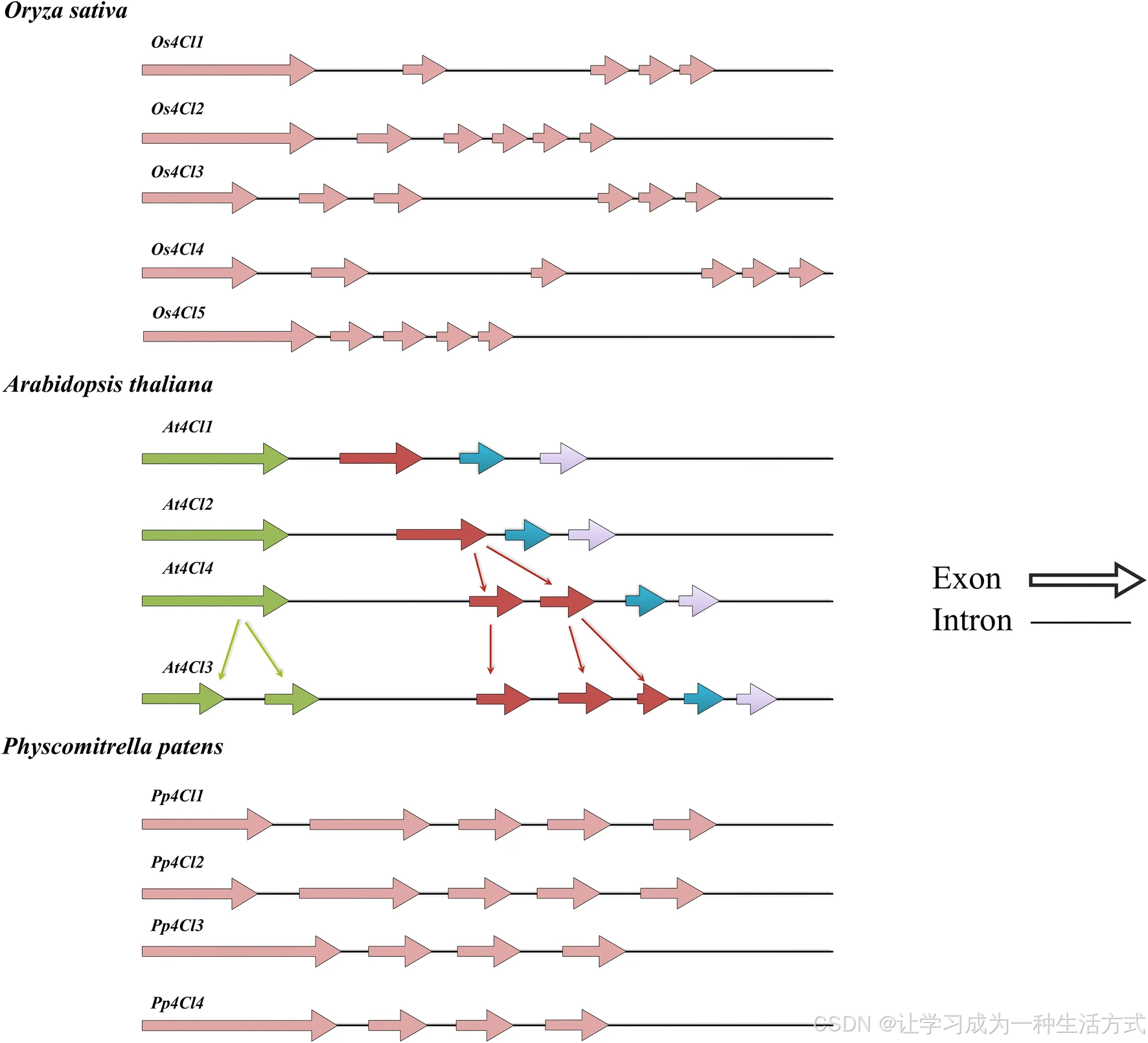
水稻(粳稻)、拟南芥和小立碗藓中 4CL 基因的结构 不同 4CL 同工型的基因结构显示,其外显子和内含子数量存在差异,这是由外显子断裂和基因复制事件导致的。箭头代表 4CL 基因的外显子,线条代表内含子。4CL:4 - 香豆酸辅酶 A 连接酶
据报道,水稻中存在 5 个 4CL 基因,分别为 Os4CL1、Os4CL2、Os4CL3、Os4CL4 和 Os4CL5。序列分析表明,Os4CL1 和 Os4CL5 均含有 5 个外显子和 4 个内含子,内含子位置保守,但长度与序列存在差异。Os4CL2 多一个内含子,该内含子由打断 Os4CL1 和 Os4CL5 的第 3 个外显子产生。
在小立碗藓中,Pp4CL1 和 Pp4CL4 具有 5 个外显子和 4 个内含子;而 Pp4CL2 和 Pp4CL3 具有 4 个外显子和 3 个长度相近的内含子。对多种 4CL 同工型的序列分析表明,外显子与内含子数量的差异源于外显子断裂,而同工型数量的变异则极有可能由基因复制事件导致。
4CL 在木质化、类黄酮合成及胁迫应答中的作用
苯丙烷途径的众多产物在植物适应环境、生长发育、繁殖以及抵御生物与非生物胁迫等方面发挥关键作用。本文重点阐述 4CL 在木质素、类黄酮、苯丙烷类及其他化合物合成中的功能,及其在机械支撑与逆境防护中的贡献。
木质化作用
木质素是仅次于纤维素的第二大类生物多聚物,存在于所有维管植物的次生细胞壁中。木质素结构的复杂程度取决于其组成单体,即 H-、G-、S - 型木质素单体的比例。这些木质素单体通过苯丙烷途径中的多条支路合成。研究者利用数学模型,分析了毛果杨单一 4CL 及 4CL 复合物(Ptr4CL3-Ptr4CL5)对木质素含量的影响,包括稳态通量分布、稳健性和稳态特性等方面。结果表明,Ptr4CL3-Ptr4CL5 复合物的存在可提升 S - 型和 G - 型木质素单体合成途径的稳健性与稳定性。
在欧洲山杨中,Pt4CL1 特异地在木质化组织(如木质部)中表达,而 Pt4CL2 在茎和叶的表皮层表达。Pt4CL1 对 5 - 羟基阿魏酸活性最高,而 Pt4CL2 不能利用 5 - 羟基阿魏酸,对香豆酸的催化效率最高。这种表达的组织分区性与底物偏好性表明,Pt4CL1 和 Pt4CL2 分别参与木质素和其他苯丙烷类物质的合成。
在杂交杨中,通过快速蛋白液相色谱检测到 3 个 4CL 蛋白同工型,它们均能催化羟基肉桂酸,但对芥子酸无活性。其中 4CL1 主要在老叶、绿茎和木质部表达,4CL2 在幼叶中表达。在毛白杨中鉴定出 5 个 Pto4CL 同工型,它们具有不同的底物特异性与催化速率,均不能以芥子酸为底物。这 5 个同工型的过表达均能显著提高木质素含量;其中 Pto4CL4 在烟草中过表达还可导致柚皮素含量上升。
从刺槐的发育木质部提取的粗蛋白中存在 3 个 4CL 同工型。Rp4CL1 偏好以对香豆酸为底物,但不能利用阿魏酸和芥子酸;Rp4CL2 和 Rp4CL3 可活化芥子酸,且对咖啡酸和对香豆酸也具有高活性。枝条粗提物也表现出相似的底物偏好模式,表明可活化芥子酸的 Rp4CL 同工型在木质化细胞中组成型表达。
类黄酮生物合成
植物可合成类黄酮、羟基肉桂酸及其衍生物,这些物质在植物中发挥多种作用,包括吸引传粉者、抵御植食动物、参与共生与化感互作,并对紫外线辐射具有重要防护作用。这类物质主要分布于叶片、茎、顶端分生组织和花粉的表皮细胞中。具有共轭双键体系的类黄酮抗氧化能力强于单共轭双键结构。它们还可通过甲基化、糖基化等修饰改变反应活性、溶解性与稳定性。此外,类黄酮可通过淬灭单线态氧、抑制活性氧(ROS)生成相关酶(环氧合酶、脂氧合酶、单加氧酶、黄嘌呤氧化酶)、金属离子螯合、自由基清除及循环其他抗氧化剂等方式抵御 ROS 损伤。各类紫外线均可诱导保护性类黄酮的合成,其中 UVC 和 UVB 的诱导作用强于 UVA。
4CL 的催化产物是类黄酮合成的必需前体。对香豆酰辅酶 A / 肉桂酰辅酶 A 与 3 分子丙二酰辅酶 A 缩合生成查尔酮,该反应由查尔酮合成酶(CHS)催化;随后查尔酮在查尔酮异构酶(CHI)作用下异构为黄烷酮。黄烷酮进一步通过多个分支途径合成各类类黄酮,包括橙酮、二氢查耳酮、黄烷酮醇、异黄酮、黄酮、黄酮醇、无色花色素、花色素和原花色素等。
已有研究发现特定的 4CL 同工型负责调控代谢通量向类黄酮合成支路分流。在拟南芥中,At4CL3 特异地在叶片、花等受光组织中表达,且与这些组织的类黄酮含量呈正相关。水稻 Os4CL2 基因也表现出类似的表达模式,该基因特异地在花药中表达,并受紫外线照射显著上调,提示其参与类黄酮合成的通量调控。
野葛是用于预防偏头痛、高血压、酒精中毒和心血管疾病的药用植物,其主要活性成分为葛根素、大豆苷、染料木苷等异黄酮类物质。该植物中 Pl4CL 在根组织中表达量最高,葛根素含量也在根中最高。经茉莉酸甲酯(MeJA)处理后,Pl4CL1 表达上调,葛根素含量提高 3 倍以上,表明 Pl4CL1 参与野葛异黄酮的生物合成。
抵御生物与非生物胁迫
多项研究表明,4CL 的表达水平在生物与非生物胁迫下显著上升,主要通过调控木质素、类黄酮及其他次生代谢物含量实现逆境防护。在拟南芥中,研究了 At4CL 对多种胁迫的响应:机械损伤可使 At4CL1 和 At4CL2 表达上调,而 At4CL3 表达下降。At4CL3 参与类黄酮合成,At4CL1 和 At4CL2 参与木质素合成。
在罗勒中,某一 Ob4CL 同工型在干旱胁迫下表达下调,这可能与其参与木质素以外的代谢物合成有关。灰杨是耐盐杨树物种,而毛果杨对盐敏感。在盐胁迫条件下培养愈伤组织发现,耐盐物种中 Pp4CL2、Pp4CL11 和 Pp4CL12 的表达被显著诱导,明显高于盐敏感物种。番茄在感染早疫病菌后,4CL 基因的转录水平显著上调。以上结果表明,4CL 在植物应对生物与非生物胁迫过程中发挥重要作用。
4CL 的活性与表达在多个水平受到严格调控
转录水平调控
苯丙烷途径相关酶的转录调控是代谢通路的核心调控方式。对赤桉 Ec4CL1 启动子区的研究发现,其 1127 bp 的 5′上游序列包含多种顺式调控与顺式作用元件,包括光响应元件、低温响应元件、脱落酸响应元件、真菌诱导子响应元件、分生组织特异性激活元件,以及限定在木质部维管组织表达的元件。
在马铃薯中研究了 St4CL1 和 St4CL1a 基因启动子上游的甲基化模式,结果显示,经致病疫霉培养滤液诱导与未诱导组的甲基化状态无明显差异。欧芹 Pc4CL 启动子在紫外线处理下会发生甲基化模式改变,但马铃薯 St4CL 启动子未出现类似变化。这种表达与甲基化差异可能是因为 St4CL 启动子缺少 Pc4CL 中位于转录起始位点 - 57 和 - 127 附近的两个保守基序。
转录因子 MYB 家族是苯丙烷途径(包括 4CL)的重要调控因子。AtMYB4 在烟草中过表达会降低 C4H、4CL1 和 CAD 基因的基础转录水平;而拟南芥 AtMYB4 突变体对 UV-B 的耐受性显著高于野生型。UV-B 照射和机械损伤可抑制 AtMYB4 表达,从而解除对 C4H 的抑制,使保护性芥子酸酯大量合成。过表达 AtMYB4 的拟南芥在 UV-B 照射下更敏感,植株死亡率达 37%,而野生型无死亡;同时这些株系芥子酸酯含量下降,类黄酮组成不变,表明 AtMYB4 负调控紫外线保护性代谢物的合成。
在甘薯中,IbMYB1a 转录因子可与 4CL 一起调控花青素合成基因。IbMYB1a 对多个花青素合成基因具有正向调控作用,这一点已通过在烟草中不同启动子驱动下过表达 IbMYB1a 得到验证:转基因株系中 PAL、C4H、4CL、CHS、CHI、F3H、DFR 和 ANS 等基因的表达均显著上升。
|------------------------------|-----------------------------------------------------------|------------------------------------------------------------------------------------|------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------|
| Gene | Factor/element | Effect | References |
| Ip4CL | IbMYB1a | Overexpression leads to anthocyanin accumulation | An et al. ([2015](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Os4CL2 | Wounding | Downregulation | Sun et al. ([2013](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Os4CL2 | UV radiation | Upregulation | |
| Pa4CL | SA and MeJA | Upregulation | Gao et al. ([2015](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Pa4CL | ABA | Downregulation | |
| Hc4CL | SA and MeJA | Downregulation | Choudhary et al. ([2013](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Hc4CL | ABA | Upregulation | |
| At4Cl1, At4Cl2 | Wound and MeJA | Upregulation | Lee and Douglas ([1996](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| At4Cl1, At4Cl2 | Peronospora parasitica infection and wounding | Upregulation | Ehlting et al. ([1999](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| At4CL3 | Peronospora parasitica infection and wounding | No change | Ehlting et al. ([1999](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Ob4Cl | Drought stress | Downregulation | Mandoulakani et al. ([2017](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Pp4CL | ABA, MeJA and GA | Upregulation | Peng et al. ([2016](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Pe4CL2, Pe4CL11, Pe4CL12 | NaCl stress | Change depends upon genotypes | Zhang et al. ([2015](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Pe4CL5 | NaCl stress | Downregulation | Zhang et al. ([2015](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Pe4CL9 , Pe4CL10 | NaCl stress | No change | Zhang et al. ([2015](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Ma4CL3 | Wounding, salicylic acid, and ultraviolet treatments | Upregulation | Wang et al. ([2016](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Mt4CL | Aluminum stress | Upregulation | Chandran et al. ([2008](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Cs4CL | Catechin treatment | Downregulation | Rani et al. ([2009](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Cs4CL | Drought stress, ABA and GA3 | Downregulation, decrease in catechin content | |
| Cs4CL | Wounding | Upregulation, increase in catechin content | |
| Sa4CL3 | Light exposure | Upregulation | Gaid et al. ([2011](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| St4CLs a | Phytophthora infestans infection and arachidonic acid | Upregulation | Fritzemeier et al. ([1987](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Ps4CLs a | Phytophthora megasperma glycinea infection | High expression in epidermal cells, oil-duct epithelial cells and developing xylem | Schmelzer et al. ([1989](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
| Pv4CL | Sclerotinia sclerotiorum | Upregulation | Oliveira et al. ([2015](#Gene Factor/element Effect References Ip4CL IbMYB1a Overexpression leads to anthocyanin accumulation An et al. (2015) Os4CL2 Wounding Downregulation Sun et al. (2013) UV radiation Upregulation Pa4CL SA and MeJA Upregulation Gao et al. (2015) ABA Downregulation Hc4CL SA and MeJA Downregulation Choudhary et al. (2013) ABA Upregulation At4Cl1, At4Cl2 Wound and MeJA Upregulation Lee and Douglas (1996) Peronospora parasitica infection and wounding Upregulation Ehlting et al. (1999) At4CL3 Peronospora parasitica infection and wounding No change Ehlting et al. (1999) Ob4Cl Drought stress Downregulation Mandoulakani et al. (2017) Pp4CL ABA, MeJA and GA Upregulation Peng et al. (2016) Pe4CL2, Pe4CL11, Pe4CL12 NaCl stress Change depends upon genotypes Zhang et al. (2015) Pe4CL5 NaCl stress Downregulation Zhang et al. (2015) Pe4CL9, Pe4CL10 NaCl stress No change Zhang et al. (2015) Ma4CL3 Wounding, salicylic acid, and ultraviolet treatments Upregulation Wang et al. (2016) Mt4CL Aluminum stress Upregulation Chandran et al. (2008) Cs4CL Catechin treatment Downregulation Rani et al. (2009) Drought stress, ABA and GA3 Downregulation, decrease in catechin content Wounding Upregulation, increase in catechin content Sa4CL3 Light exposure Upregulation Gaid et al. (2011) St4CLs a Phytophthora infestans infection and arachidonic acid Upregulation Fritzemeier et al. (1987) Ps4CLs a Phytophthora megasperma glycinea infection High expression in epidermal cells, oil-duct epithelial cells and developing xylem Schmelzer et al. (1989) Pv4CL Sclerotinia sclerotiorum Upregulation Oliveira et al. (2015))) |
研究者将全长及截短的 4CL 启动子 与 GUS(β- 葡萄糖苷酸酶)基因融合,探究了 4CL 基因的发育阶段特异性表达与损伤诱导表达 。在转 4CL 启动子(全长 / 截短)驱动 GUS 的拟南芥中:
- GUS::At4CL1 和 GUS::At4CL2 仅在根和地上器官的维管组织 中表达;
- At4CL3::GUS 在非维管组织(叶片、子叶)、下胚轴上部和根中表达量更高;
- At4CL4::GUS 则仅在根中 表达。
调控元件分析显示:
- At4CL2 启动子 −950 ~ −750 bp 区域调控早期损伤响应 ;
- −950 ~ −1600 bp 区域对早期损伤响应起负调控 作用。At4CL2 的晚期损伤响应 可能源于其第 1/2/3 内含子中存在晚期损伤响应元件。包含全部 3 个内含子的构建体在损伤处理 72 小时后表现出强烈响应,显著强于内含子较少或无内含子的构建体。
植物激素对 4CL 的调控
植物激素通过调控 4CL 表达影响包括苯丙烷途径在内的多条代谢通路(表 2)。例如,用脱落酸(ABA)、水杨酸(SA)和茉莉酸甲酯(MeJA)处理 2 月龄地钱叶状体,结果显示:SA 和 MeJA 处理使Pa4CL1 上调,而 ABA 处理使其下调。红麻 Hc4CL 在 MeJA 和 SA 处理下表达下调;但在 ABA 处理下,转录水平先在 1 小时轻微下降,随后持续上升并在 24 小时达到峰值。烟草 Nt4CL1 和 Nt4CL2 的表达可被机械损伤和 MeJA 诱导。在丹参中,SmC4H1、Sm4CL2、Sm4CL3 及多个 Sm4CL-like 基因的启动子区含有 MeJA 响应元件,其表达可能受 MeJA 调控。
拟南芥、象草、川桑、茶树等植物的 4CL 基因激素调控信息见表 2。不同植物中特定 4CL 同工型对同一激素的响应存在明显差异:例如 SA 处理使地钱 Pa4CL 上调,却使红麻 Hc4CL 下调,表明各 4CL 同工型对胁迫信号具有差异化响应 。
生物与非生物胁迫下的表达调控
多项研究表明,4CL 基因表达受干旱、盐胁迫、温度等多种非生物胁迫调控(表 2)。在拟南芥中,机械损伤后:
- At4CL1、At4CL2 在 2.5 小时表达上升,随后回落,48 小时再次达到峰值;
- At4CL3 表达先下调,4 小时恢复正常,之后持续上升至 72 小时;
- At4CL4 在 2.5 小时内上升并维持高表达至 12 小时,随后回落至基础水平。
不同研究得到的 At4CL3 表达模式略有差异。
在粳稻中,损伤处理显著上调 Os4CL3、Os4CL4、Os4CL5 及其他苯丙烷途径基因,而 Os4CL1、Os4CL2 下调。经暗适应的水稻植株在 UV 照射下,Os4CL1、3、4、5 均下调,仅 Os4CL2 上调,提示 Os4CL2 很可能参与黄酮醇合成。
在干旱胁迫下,罗勒 Ob4CL 与 ObC4H 表达下降,CVOMT、EOMT 上升,CAD 相对稳定,伴随草蒿脑、甲基丁香酚等精油成分含量升高,表明干旱诱导的基因表达改变直接导致代谢谱重塑。
盐胁迫(NaCl 处理)可显著上调 PAL、CCoAOMT、C3H、HCT、F5H 等木质素合成相关基因。胡杨、川桑、蒺藜苜蓿、茶树、花楸等植物中的研究也证实 4CL 在非生物胁迫应答中发挥重要作用(表 2)。
植物可通过调控苯丙烷途径抵御细菌、真菌、病毒侵染及昆虫取食等生物胁迫。作为关键酶,4CL 的表达在生物胁迫下显著改变(表 2),表明其在抗病抗虫中具有重要作用。马铃薯叶片受致病疫霉侵染后,PAL、4CL 等苯丙烷途径基因快速积累,侵染部位迅速褐变并发生超敏反应。侵染 2 小时后 PAL 和 4CL 的 mRNA 翻译活性最强,6--12 小时酶活性提升约 2 倍。
拟南芥 --- 寄生霜霉、马铃薯 --- 致病疫霉、欧芹 --- 大雄疫霉、菜豆 --- 核盘菌等多种植物 --- 病原互作系统均观察到 4CL 基因上调。葡萄红斑驳病毒(GRBaV)侵染会导致果实成熟期葡萄苯丙烷途径基因(含 4CL)整体下调。
4CL 同工型的功能特异性:案例分析
利用 STRING 数据库分别对拟南芥 At4CL1、At4CL3 和大豆 Gm4CL4 进行共表达网络分析 (交互数量设为 20,其余参数默认)并整合结果(图 5)。
- At4CL1 与木质素合成基因 共表达;
- At4CL3 与类黄酮合成基因 共表达。
这表明不同 4CL 同工型负责不同支路的代谢通量分流 。At4CL1 和 At4CL3 同时也与木质素 / 类黄酮以外的其他代谢途径基因共表达。Gm4CL4 则同时与木质素、类黄酮、其他代谢物合成基因及未功能注释基因共表达。
基于拟南芥(At4CL1/At4CL3)和大豆(Gm4CL4)的共表达分析可见:
- 部分植物中存在专一同工型 ,分别负责木质素或类黄酮合成;
- 部分植物中则存在多功能同工型 ,可同时参与两条支路。
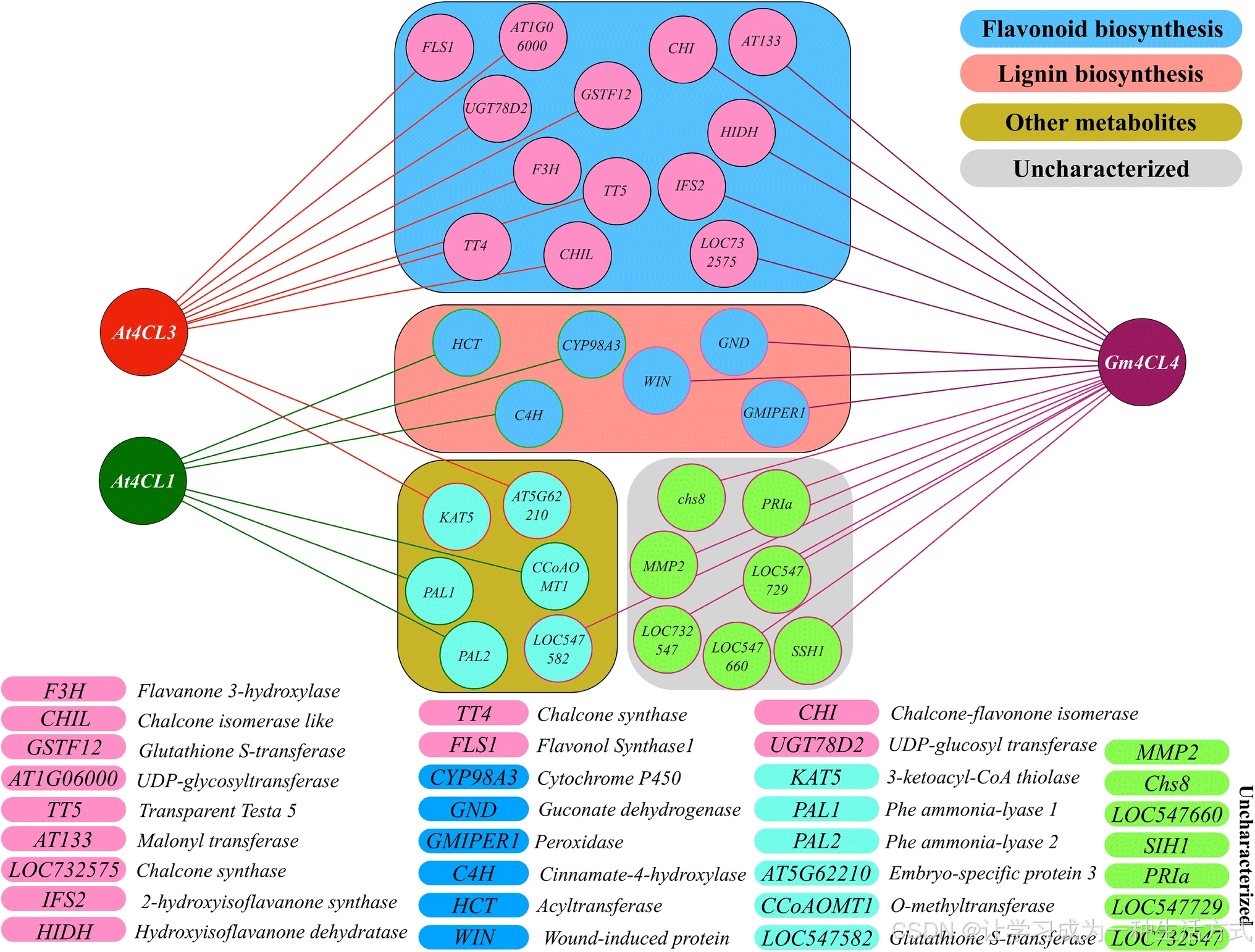
使用 STRING 数据库(相互作用基因 / 蛋白质检索工具)分别对 At4CL1、At4CL3 和 Gm4CL4 进行共表达分析,并将结果整合。At4CL1 和 At4CL3 分别与木质素生物合成和类黄酮生物合成相关基因共表达,而 Gm4CL4 同时与两个途径的相关基因共表达。除需展示的互作数量设置为 20 外,其余均采用默认参数。本图依据各 4CL 在 STRING 分析中的结果,由补充材料图 S1 整理绘制。4CL:4 - 香豆酸 - 辅酶 A 连接酶;At:拟南芥;Gm:大豆
4CL 在多种产物代谢途径工程中的应用前景
苯丙烷代谢和类黄酮代谢相关酶以膜结合多酶复合体(即代谢)的形式存在于内质网的细胞质一侧。这些有序的多酶复合体能够高效地将中间体在连续反应的酶之间进行底物通道化。代谢的相互作用与组成在不同植物物种中可能存在差异。正如 Chemler 和 Koffas(2008)综述所述,通过基因克隆并在细菌(大肠杆菌)和酵母(酿酒酵母)中表达 4CL 及其他参与其生物合成的酶,可以实现类黄酮的合成。
燃料工业
木质素与纤维素微纤丝共同为植物提供机械强度。木质素含量影响植物材料在化学品、纤维和能源生产等方面的利用效率。木质素是苯丙烷途径的产物之一,特定 4CL 同工型的表达在木质素生物合成中发挥作用。沉默 Pt4CL1 基因的转基因山杨(Populus tremuloides Michx.)表现出木质素生物合成下调。这类植株的木质素最多可降低 45%,而纤维素含量增加 15%。沉默株系的茎更粗、节间更长、叶片更大,整体生长速率高于对照植株。甘蔗是生产生物乙醇的最佳原料。从甘蔗杂交种中鉴定出两种 4CL 同工型。基于系统发育分析和 RNAi 沉默结果,其中 Sh4CL1 参与木质素生物合成。RNAi 抑制 Sh4CL1 可使木质素含量降低 16.5%,与野生型对照相比,糖化效率提升 52--76%。这表明 4CL 沉默可用于改良木质纤维素原料,以用于生物燃料生产。
天然产物工业
苯丙烷通用途径可产生多种具有重要价值的天然产物(NPs)。利用 4CL 生产高价值天然产物已在细菌、真菌和植物系统中得到验证。白藜芦醇是一种重要的次生代谢产物,具有保健功能,存在于红酒中。将拟南芥 At4CL1 与葡萄芪合酶(VvSTS)融合,可用于在酵母中提高白藜芦醇的合成。使用该融合蛋白(At4CL1-VsSTS),白藜芦醇的含量相较于分别表达这些酶的酵母提高了 15 倍。迷迭香酸也是苯丙烷途径的重要产物之一,由 4 - 羟基苯乳酸与 4 - 香豆酰辅酶 A 缩合生成,该反应由羟基肉桂酰辅酶 A: 羟基肉桂酰转移酶催化。它具有多种生物活性,包括抗病毒、抗菌、抗氧化和抗炎活性。茉莉酸甲酯(MeJA)处理藿香(Agastache rugosa Kuntze)细胞培养物可使迷迭香酸积累量提高 2.1 至 3.9 倍。与未处理的培养细胞相比,ArPAL、Ar4CL 和 ArC4H 的转录水平分别提高了 4.5 倍、3.4 倍和 3.5 倍。姜目植物可产生姜黄素类化合物,具有广泛用途,包括食品添加剂、兴奋剂、食品着色剂、抗肿瘤、抗氧化和保肝活性等。通过在大肠杆菌中克隆紫草 4CL 和水稻姜黄素合成酶(CUS),实现了姜黄素类化合物的生物合成。在表达 4CL 和 CUS 的重组大肠杆菌培养过程中,使用米糠油生产工业废弃物米糠沥青中的阿魏酸生产姜黄素类化合物。圣罗勒中的 Os4CL 负责将代谢通量分配至丁香酚生物合成。丁香酚具有抑制糖尿病晚期糖基化终产物的作用。4CL 在提升天然产物合成方面具有突出作用。
食品技术
研究了苹果果实成熟过程中参与木质素单体和类黄酮生物合成基因的表达模式。PAL 和 4CL 同工型的差异表达导致果实成熟过程中木质素生物合成受到抑制,而类黄酮生物合成被强烈诱导。结果表明,通过 PAL 和 4CL 等分支点酶同工型的差异表达可实现途径的精细调控。在梨果实发育的不同阶段观察到特定 4CL 同工型的差异表达。因此,利用对 4CL 的操控进行代谢途径工程,在采后处理和食品技术相关产业中具有应用价值。
结论与未来展望
4CL 本质上参与调控不同苯丙烷类化合物生物合成的前体分配。基于系统发育分析,4CL 分为四个类群,即参与木质素单体生物合成的 I 型、参与非木质素类苯丙烷化合物生物合成的 II 型,以及存在于单子叶植物中、功能分别与 I 型和 II 型相似的 III 型和 IV 型。单子叶植物和双子叶植物的 4CL 基因在物种分化后独立进化。然而,I 型和 II 型可能在裸子植物与被子植物谱系分化之前就已经分化。4CL 的活性和表达在时空上受到严格调控,同时也响应生物和非生物胁迫。
借助生物信息学工具、晶体学和定点突变等技术,4CL 底物结合口袋(SBP)氨基酸及结构域的功能已得到深入研究。通过定点突变和结构域交换方法已成功提高其活性。
对多种植物中已功能鉴定的 4CL 进行共表达分析表明,部分植物仅需一个 4CL 即可同时参与木质素和类黄酮生物合成,而另一些植物则拥有分别负责这两个生物合成途径的不同 4CL。
代谢途径在其终产物生物合成的起始点、分支点和 / 或某些情况下由中间酶进行调控。然而,大多数代谢途径中研究较多的是末端酶。目前对 4CL 的研究大多集中在功能鉴定和外界因子调控方面。内源因子 / 信号对 4CL 功能和调控的影响仍不明确,而这是挖掘其在代谢工程中进一步应用的重要方面。已有研究证实,特定 PAL 同工型在苯丙烷途径调控中也发挥重要作用,PAL 是该途径的第一个关键步骤。
为实现高效的代谢途径工程,详尽的验证是必要的:(a) 中间代谢物和终产物对特定 4CL 同工型活性的影响;(b) 转录因子在调控各 4CL 同工型水平中的作用;(c) 与其他途径的交叉互作;(d) 形成多种同工型的剪接机制;(e) 翻译后水平的调控;(f) 在细菌或酵母宿主系统中的活性优化等。此类信息对于实现具有商业价值的目标代谢物的高效代谢途径工程至关重要。
4CL:4 - 香豆酸:辅酶 A 连接酶C3H:对香豆酸 3 - 羟化酶C4H:肉桂酸 4 - 羟化酶CAD:肉桂醇脱氢酶CCoAOMT:咖啡酰辅酶 A O - 甲基转移酶PAL:苯丙氨酸解氨酶CVOMT:胡椒酚 O - 甲基转移酶COMT:咖啡酸 O - 甲基转移酶EOMT:丁香酚 O - 甲基转移酶F5H:阿魏酸 5 - 羟化酶CHS:查尔酮合成酶ANS:花青素合成酶HCT:对羟基肉桂酰辅酶 A:奎宁酸 / 莽草酸对羟基肉桂酰转移酶DFR:二氢黄酮醇 4 - 还原酶。