本笔记基于杨荣武教授主编的《生物化学》第四版,配图也源于教材
(这里说的自由能是很大的负值指的都是自由能是负值且其绝对值很大)
三羧酸循环也称为柠檬酸循环或者Krebs循环,可以分为氧化型和还原型两种形式,我们主要介绍氧化型TCA循环。
这是完整的TCA循环,在我们分别讲述各步酶促反应之后,将汇总再次叙述
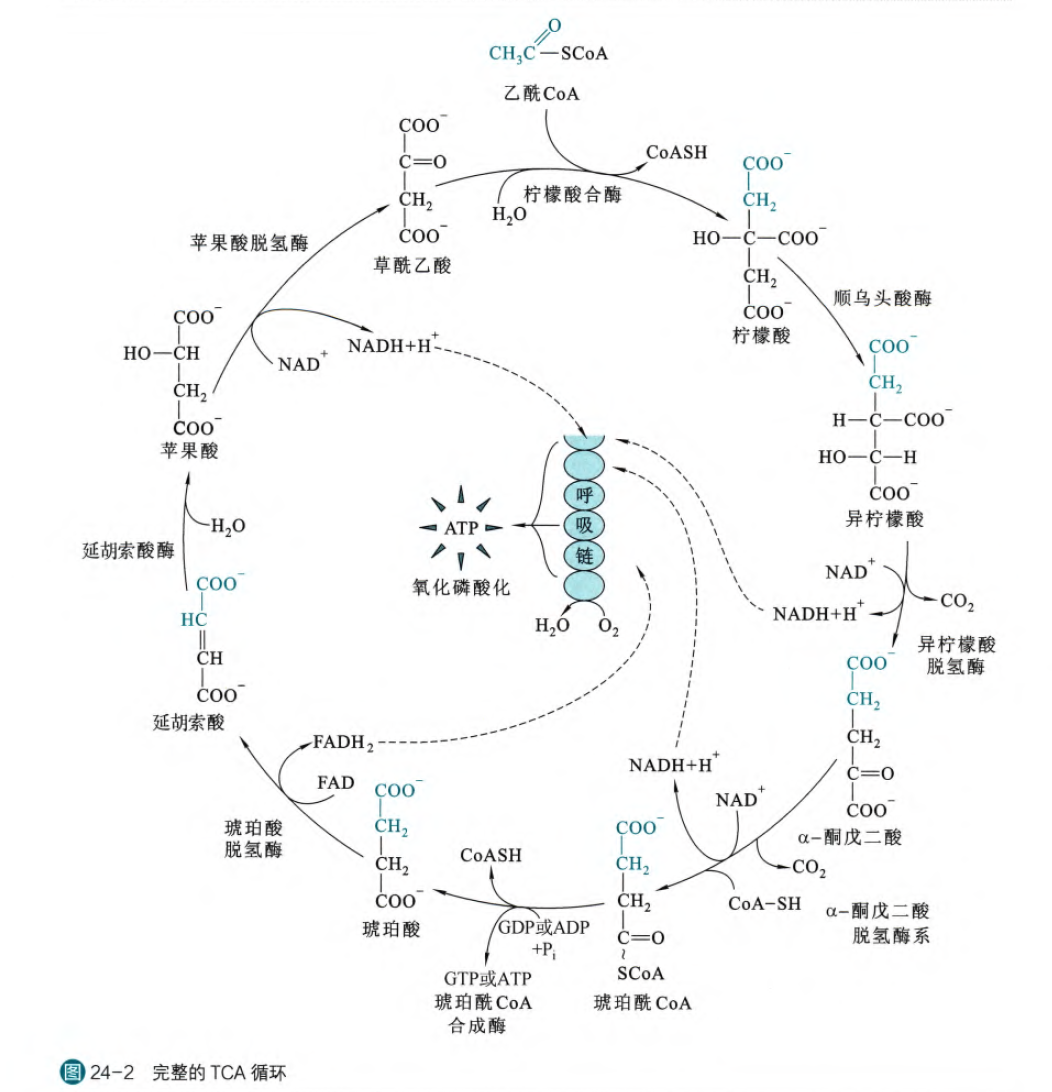


- 柠檬酸的合成:

这步反应是由柠檬酸合酶催化发生的克莱森酯缩合反应,这步反应的自由能是一个很大的负值,所以是不可逆反应。这一步反应实际上经历了一个称为柠檬酰CoA的中间物,而且这一个酶促反应也完美体现了诱导契合模型(在没有底物结合的时候,两个构成柠檬酸合酶的亚基呈现开放状态,在底物结合之后,两个亚基变为封闭状态);首先草酰乙酸进入酶结合位点,诱导酶结构发生改变之后,暴露出乙酰CoA的结合位点,当乙酰CoA结合之后,缩合形成柠檬酰CoA,形成柠檬酰CoA之后,进一步诱导远离酶活性中心的Asp拉到柠檬酰CoA的硫酯键附近,发生催化,诱导其断裂,之后产物被依次释放(反应动力学典型的序列有序型)
那么这样一个反应其实类似于激酶,避免乙酰CoA不当水解。
那么乙酸的类似物------氟代乙酸可以强烈地抑制TCA循环,氟代乙酸CoA可以与草酰乙酸反应生成氟代柠檬酸。然后氟代柠檬酸会在顺乌头酸酶的催化下,首先变为氟代顺乌头酸,然后再添加氢氧根离子之后,丢掉氟原子,变为4-羟基反乌头酸,这种物质可以紧密结合在顺乌头酸的活性中心,导致酶活的抑制(那么这一种底物类似物抑制剂在糖酵解当中也出现了,即2-脱氧葡萄糖,可以与己糖激酶结合被磷酸化,但是不可以被变构所以被积累并反馈抑制葡萄糖磷酸化过程)
- 异柠檬酸的形成
这一步是将柠檬酸异构成异柠檬酸的反应,之所以这个反应是必要的是因为柠檬酸不是一个很好的氧化反应底物,但是经过异构之后得到的异柠檬酸将原本不易氧化的三级羟基变为了易氧化的二级羟基。
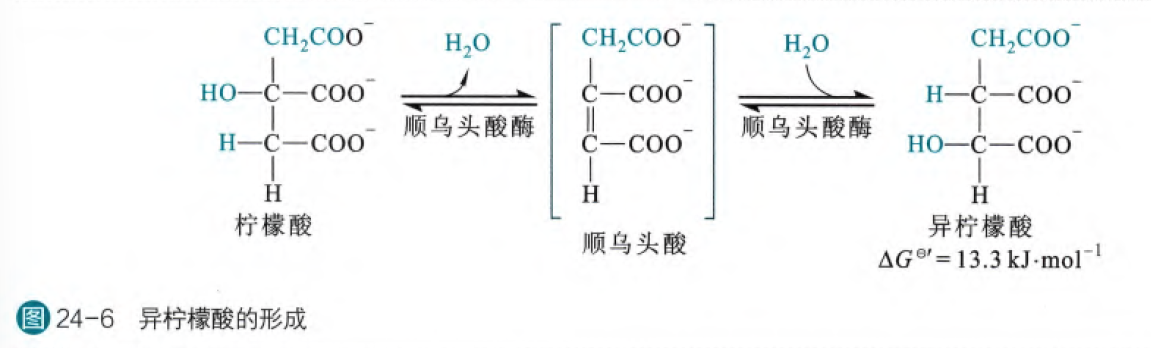
这个反应由顺乌头酸酶催化,首先柠檬酸脱水生成顺乌头酸,然后再加上一分子水变为异柠檬酸,这个反应虽然是自由能增大的反应,最后柠檬酸:顺乌头酸:异柠檬酸的比例=90:4:6,但是由于下一步的不可逆反应的拉动,使得异柠檬酸可以不断地生成
顺乌头酸酶以铁硫蛋白作为辅因子,其中的铁硫簇直接参与催化反应。铁硫蛋白不仅可以作为TCA循环的酶之一,同时也可以作为细胞内铁离子的感应器,参与铁蛋白和转铁蛋白受体在翻译水平上的表现
- 异柠檬酸的氧化脱羧
这一步氧化脱羧反应由异柠檬酸脱氢酶(IDH)催化,反应分为两步,首先发生脱氢反应生成草酰琥珀酸,然后再发生β脱羧反应,生成α-酮戊二酸,这个反应的自由能是一个很大的负值,所以是不可逆反应(虽然一般的脱氢反应是可逆的,但是脱羧反应是不可逆的,所以这个反应也是不可逆的)
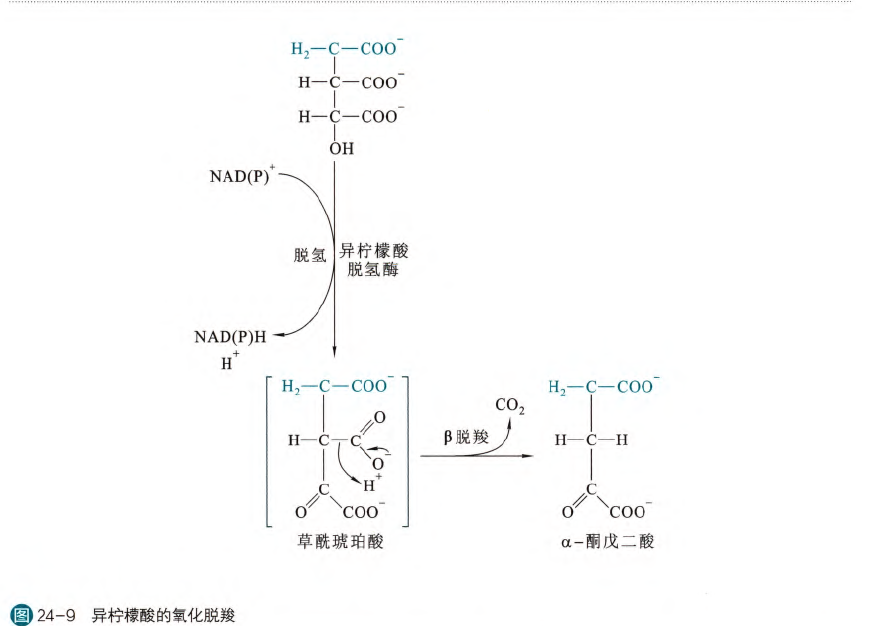
目前真核细胞内发现了三种异柠檬酸脱氢酶------IDH1,IDH2和IDH3,都是由核基因编码。其中IDH1和IDH2用辅酶II作为辅因子,而IDH3用辅酶I作为辅因子。IDH1存在于细胞质基质和过氧化物酶体当中,而IDH2和IDH3仅仅存在于线粒体基质中。参与TCA循环的主要是IDH3,释放出的NADH直接在复合体I的作用下脱氢。而IDH1和IDH2的作用主要是在细胞缺氧的时候将谷氨酰胺衍生的α-酮戊二酸羧化再还原为异柠檬酸。异柠檬酸转变为柠檬酸之后再被裂解为草酰乙酸和乙酰CoA
- α-酮戊二酸的氧化脱羧
这一步与丙酮酸的氧化脱酸很相似,涉及的反应机理也基本相同,关键在于底物的不同。
α-酮戊二酸脱氢酶系也有三个酶组成:α-酮戊二酸脱氢酶、二氢硫辛酸转琥珀酰酶和二氢硫辛酸脱氢酶,对于前面两个酶,其差别其实在于催化的底物不同,特别是对于E2,丙酮酸的是二氢硫辛酸转乙酰酶,但是对于第三个酶即是完全相同的啦
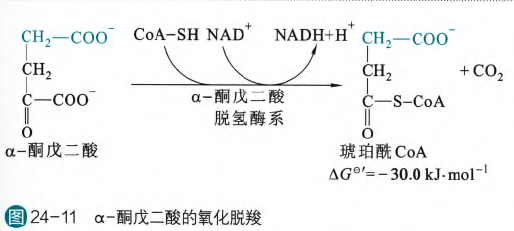
这一步也是很大的负值,在胞内也是不可逆反应
- 底物水平的磷酸化
这是TCA循环内唯一一部的底物水平磷酸化,由琥珀酸CoA合成酶催化,这个酶也是根据逆反应的性质而定义的
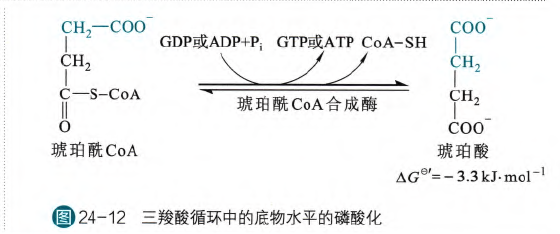
在这个反应过程当中,首先磷酸基团会亲核进攻羰基碳,形成琥珀酰磷酸,然后酶活性中心的His的咪唑基的N会亲核进攻羰基碳,形成琥珀酸,同时磷酸连接在His上,形成磷酸化的His,之后His可以把无机磷酸转移给GDP或者是ADP,形成GTP和ATP
但是究竟是生成ATP还是GTP还是要关注这个细胞对于ATP和GTP的需求,比如对于高耗能的心肌细胞,主要通过ATP来实现肌肉收缩,所以主要使用ATP专一的琥珀酰CoA合成酶;对于合成代谢非常旺盛的肝细胞,主要是使用GTP专一的琥珀酰CoA合成酶
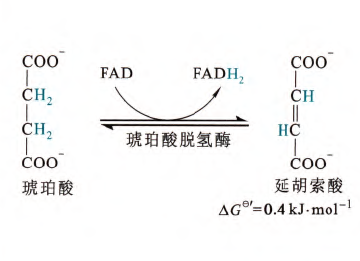
- 琥珀酸的脱氢
这是TCA循环中的第三次脱氢反应,有琥珀酸脱氢酶催化,产物是反丁烯二酸(也称为延胡索酸和富马酸),琥珀酸的类似物丙二酸是该酶的竞争性抑制剂(琥珀酸脱氢酶也属于呼吸链复合体II)
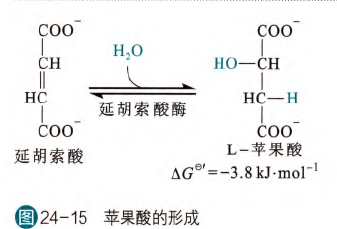
- 苹果酸的形成
这是一步水合反应,由延胡索酸催化,该酶经常被称为延胡索酸水合酶(FH)或者富马酸酶
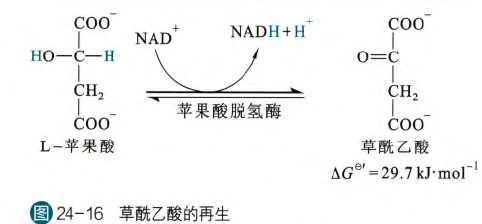
- 草酰乙酸的再生
这是TCA循环的最后一步,也是TCA循环的第四次脱氢反应,由苹果酸脱氢酶催化,在胞内的自由能变化接近于0,而且在第一步的草酰乙酸和CoA的反应的拉动下,可以不断反应
三羧酸循环总结:
-
在整个三羧酸循环中,一个出现3步不可逆反应,第一步是柠檬酸合酶催化的草酰乙酸转变为柠檬酸的反应,第二步是异柠檬酸脱氢脱羧转变为α-酮戊二酸的反应,第三步是α-酮戊二酸经过α-酮戊二酸脱氢酶系转变为琥珀酰CoA的反应,其他的反应都是自由能差异较小的可逆反应,同时我们也可以理解为这些可逆反应是被这三步不可逆反应拉着前进的。
-
在整个三羧酸过程当中,只有异柠檬酸和苹果酸是具备有手性的,对于顺乌头酸酶是具备立体专一性的,但是因为延胡索酸是对称的,所以延胡索酸酶的立体专一性体现不了
-
一共存在4步脱氢反应和2步脱羧反应,其中两步脱羧反应都是和脱氢反应相耦合的。这两部反应分别是异柠檬酸脱氢酶(由6碳->5碳)和α-酮戊二酸脱氢酶系(5碳->4碳)催化的,这两步也都是不可逆反应,另外两步的脱氢反应分别是琥珀酸脱氢酶和苹果酸脱氢酶,其中琥珀酸脱氢酶是产生了FADH2而不是NADH,这个酶也是复合体II的重要单位
-
这个循环仅仅涉及一步底物水平磷酸化,也就是琥珀酰CoA经过琥珀酰CoA合成酶催化变为琥珀酸的反应,这一步可以产生ATP,也可以产生GTP,至于产生哪一种还是需要根据细胞对两种能量货币的需求不同来决定
-
这个反应一共涉及到水分子参与反应的有三步,首先是柠檬酸合成过程当中,形成的柠檬酰CoA的高能硫酯键的断裂是需要水分子的加入,第二步是顺乌头酸酶催化的反应当中首先需要脱水生成顺乌头酸然后加水生成异柠檬酸,但是总体上不涉及水分子的消耗,第三步是延胡索酸酶催化的苹果酸的合成,这一步需要水分子的参与
-
对于质子的生成与消耗,首先在生成NADH的反应当中,每生成一分子NADH,都会生成一个质子,同时因为在第一步的柠檬酸生成的过程当中,柠檬酸水解释放一个质子,所以总反应一共生成4个质子,然后在FADH2合成的时候需要消耗两个质子,所以总共一个三羧酸循环结束生成2个质子
-
在整个循环更多当中,可以理解为一直在氧化释放乙酰基的能量,但是实际上本轮三羧酸循环羧化和脱氢的对象并不是乙酰基。可以看到,在异柠檬酸脱氢酶和α-酮戊二酸脱氢酶系的两步脱羧反应脱下的碳都是原本草酰乙酸上已经羧化的碳,而乙酰基加入的碳一个变为羧基碳在下一轮即会被脱去,而另一个碳变为了羰基碳,在下下轮循环才会被脱去,所以被同位素标记的乙酰辅酶A的碳在经过一轮循环之后才可以在CO2检测到
-
虽然在这个过程当中并不涉及氧气,但是氧气对于NADH和FADH2的消耗是至关重要的。
-
在这个过程当中,一个乙酰辅酶A可以生成3个NADH,1个FADH2和1个ATP/GTP,如果经过呼吸链的NADH和FADH2的代换,那么可以生成10个ATP,所以一个葡萄糖经过糖酵解可以生成2个ATP和2个NADH,经过换算得到7个ATP,丙酮酸脱氢酶系可以生成2个NADH,所以最佳情况可以生成32个ATP
(也就是一个葡萄糖在糖酵解可以产生2个NADH和2个ATP,但是因为NADH是在细胞质基质当中产生的,所以由细胞质基质运输到线粒体基质呼吸链复合体I时,如果经过GPS进入的话,每个NADH仅能转化1.5个ATP,如果经过MAS进入的话,那么可以转化为2.5个ATP,所以糖酵解可以产生5个或者6个或者7个ATP,再加上丙酮酸脱氢酶系得到的2个NADH(在线粒体基质,所以是转化为5个ATP),最后加上三羧酸循环产生的20个ATP,最终可以得到30/31/32个ATP)
三羧酸循环的功能
-
三羧酸循环产生的ATP远远高于糖酵解产生的ATP,具体数量在上面已经分析
-
很多中间代谢物可以离开:
(1)柠檬酸:柠檬酸可以通过线粒体内膜上的柠檬酸转运蛋白运输到细胞质基质,然后一方面可以在ATP柠檬酸裂合酶的催化下,可以裂解为草酰乙酸和乙酰CoA,其中乙酰CoA既可以作为脂肪酸和胆固醇的原料,又是蛋白质乙酰化修饰的乙酰基供体,另一方面还可以作为PFK1的别构抑制剂(果糖-1-磷酸激酶,可以催化其转变为果糖-1,6-二磷酸),抑制糖酵解的活性,或者作为果糖-1,6-二磷酸酶的别构激活剂,激活糖异生,或者作为乙酰CoA的羧化酶的别构激活剂,刺激脂肪酸的生物合成。
(2)顺乌头酸:可以在顺乌头酸脱羧酶的催化下脱羧,转变为衣康酸,衣康酸可以参加很多生物过程,比如参与免疫调节
(3)α-酮戊二酸:既可以作为底物,经过转氨基反应转变为谷氨酸,还可以作为辅助底物,参与多种依赖氧气的有羟化酶催化的氧化反应,比如胶原蛋白和HIFα的羟基化修饰
(4)琥珀酰CoA:可以作为血红素合成的原料,还可以参与酮体的利用;琥珀酰CoA也是许多蛋白质分子上的Lys参加发生琥珀酰化修饰的琥珀酰基的供体
(5)富马酸(延胡索酸):可以参与一些蛋白质分子上的Cys残基上的琥珀酰化修饰,在细胞缺氧的时候,富马酸可以介入并且临时充当呼吸链的末端电子受体,有助于维持缺氧时线粒体的功能
(6)草酰乙酸:可以作为糖异生的原料,还可以参加MAS的穿梭系统,还可以作为Asp和Asn合成的原料
- 产生CO2:一轮TCA可以经过两次脱羧,产生两个CO2
三羧酸循环的回补反应:
因为TCA循环当中的多种代谢中间物可以离开参与其他代谢途径,所以需要配备专门的回补反应,来补充草酰乙酸或者其前体的量,以维持TCA循环的正常速率(因为这是一个循环,所以不需要对于每一个中间物都进行回补)
TCA循环的回补反应主要在草酰乙酸、α-酮戊二酸、琥珀酰CoA和苹果酸
- 草酰乙酸的回补:
这是主要的形式,相关的酶有PEP羧化酶,丙酮酸羧化酶和PEP羧激酶
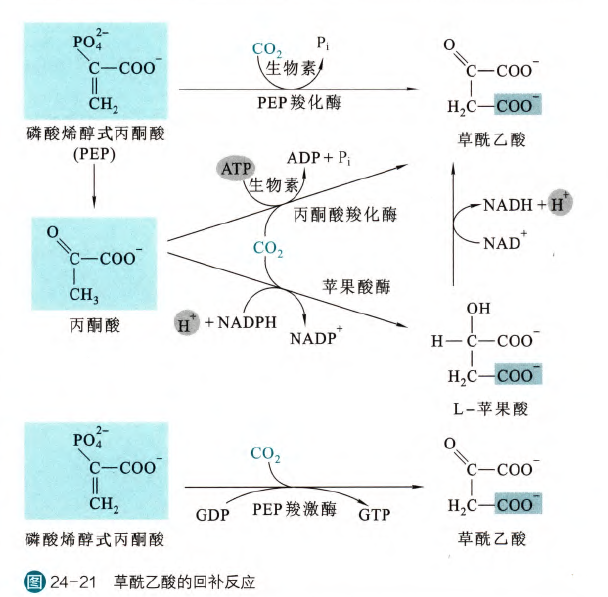
- α-酮戊二酸的回补
由谷丙转氨酶催化的转氨基反应,或者谷氨酸脱氢酶催化的氧化脱氨基反应可以将谷氨酸转化为α-酮戊二酸,且都发生在线粒体基质,因此都可以充当回补反应
- 琥珀酰CoA的回补
异亮氨酸、缬氨酸、甲硫氨酸和苏氨酸这四种氨基酸均可以被氧化为琥珀酰CoA,奇数脂肪酸在胞内氧化也可以产生琥珀酰CoA
- 苹果酸的回补
由苹果酸酶催化
三羧酸循环的调控
与糖酵解类似的,对TCA本身的调控主要集中在柠檬酸合酶、异柠檬酸脱氢酶和α-酮戊二酸脱氢酶系三个限速酶上。此外,乙酰CoA是进入TCA循环的底物,机体可以通过控制乙酰CoA的形成来控制TCA循环,所以细胞内产生CoA的酶也会参与到TCA的调控。
- 柠檬酸合酶的调控:
对柠檬酸合酶的调控主要是别构调节,细胞高能状态的指示剂(NADH、ATP),反应的中间产物(琥珀酰CoA)或者终产物都可以作为别构抑制剂来抑制酶的活性,而细胞低能状态的指示剂ADP可以作为别构激活剂来刺激酶的活性。此外,高浓度的柠檬酸可以通过竞争的方式抑制柠檬酸合酶。(在糖酵解当中,己糖激酶可以受到其产物葡萄糖-6-磷酸的抑制)
- 异柠檬酸脱氢酶的调控
对异柠檬酸脱氢酶的调控有共价修饰和别构调节。作为异柠檬酸脱氢酶的别构抑制剂主要是ATP,别构激活剂的是ADP和钙离子。同时钙离子可以作为异柠檬酸脱氢酶、α-酮戊二酸脱氢酶以及丙酮酸脱氢酶系的别构激活剂(因为钙离子是肌细胞进行收缩的信号,而收缩需要ATP)
- α-酮戊二酸脱氢酶系的调控
对α-酮戊二酸脱氢酶系的调控有别构调控和产物的竞争性反馈抑制。钙离子和ADP别构激活α-酮戊二酸脱氢酶,琥珀酰CoA和NADH分别反馈抑制二氢硫辛酸转琥珀酰酶和二氢硫辛酸脱氢酶。
α-酮戊二酸脱氢酶系可以视为线粒体内氧化还原状态的感应器,可以调节线粒体内的功能状态,防止氧化损伤。因为α-酮戊二酸可以直接产生NADH,所以可以通过调节这一步反应的调控,从而降低呼吸链的强度,减少自由基的产生。
如果线粒体内的自由基水平高时,α-酮戊二酸可以受到自由基调节的可逆抑制。这种暂时的可逆抑制来自于α-酮戊二酸脱氢酶系的E2发生的谷胱甘肽化修饰,在有自由基的时候,该酶被关闭,而一旦自由基被细胞清除,那么该酶在谷氧还蛋白作用下恢复活性。在谷胱甘肽化的修饰下,有利于避免硫辛酰胺的氧化,因为硫辛酰胺会直接影响到其他三个酶的活性,所以这样的修饰有助于避免整个酶系受到损伤。
- 丙酮酸脱氢酶系的调控
(1)竞争性反馈抑制(终产物与酶的活性中心竞争性结合):乙酰CoA和NADH可以反馈抑制E2和E3
(2)ADP、NAD+作为细胞低能态的指示剂,可以别构激活丙酮酸脱氢酶系;NADH可以别构抑制
(3)参与丙酮酸脱氢酶共价修饰的酶是丙酮酸脱氢酶激酶和磷蛋白磷酸酶,两者都与丙酮酸脱氢酶系紧密结合,激活催化特定位点的丝氨酸残基的磷酸化,导致其活性丧失;磷酸酶可以水解该磷酸修饰,使其活性恢复。激酶和磷酸酶又同时受到别构调节其中低能态指示剂(ADP和NAD+)可以别构抑制激酶活性,CoA和丙酮酸(原料)可以别构抑制激酶活性,钙离子(需能指示剂)可以别构激活磷酸酶活性,这五种物质维持丙酮酸脱氢酶的去磷酸化;相反,乙酰CoA和NADH激活激酶活性,维持丙酮酸脱氢酶的磷酸化形式
- 乙酰化修饰对TCA循环酶的调节
蛋白质的乙酰化是一种动态、可逆性的翻译后修饰,可以使得细胞感知能量状态并且对环境变化和压力做出反应。绝大多数生物体(需氧生物存在三羧酸循环的)TCA循环酶都可以发生乙酰化修饰,特别是异柠檬酸脱氢酶、琥珀酸脱氢酶和苹果酸脱氢酶,这些酶会因为乙酰化而发生抑制。SIRT3是位于线粒体的一种依赖NAD+的脱乙酰酶,因此可以通过脱乙酰化来激活这些酶。所以SIRP3水平较高的话,那么代谢一般也会越旺盛
乙醛酸循环
乙醛酸循环也称为TCA循环支路,如果是植物,那么乙醛酸循环存在于一种由膜包被的细胞器即乙醛酸循环体当中。它于TCA循环的前两步和最后一步是一样的,差别在于异柠檬酸形成之后。
在形成异柠檬酸之后,异柠檬酸会在异柠檬酸裂合酶的作用裂解为乙醛酸和琥珀酸。产生的琥珀酸离开循环,但是乙醛酸留在循环内,与第二个乙酰CoA在苹果酸合酶的作用下形成苹果酸,苹果酸在最后一步脱氢得到草酰乙酸。
所以一个乙醛酸循环,需要两个乙酰CoA,这个循环的碳不是被脱羧离去,是以琥珀酸的形式离去(4碳单位)。那么在这个过程当中,在苹果酸脱氢产生草酰乙酸的过程当中会产生1分子的NADH,所以在这个循环当中,可以简单理解为,两分子的乙酰基团转变为了一分子的琥珀酸和一分子的NADH。

乙醛酸循环的功能
乙醛酸循环的功能实际上是摆脱了糖酵解只能使用六碳单位有机物或者其中代谢途径当中的有机物作为能源物质的局限性,使得任意的能够产生二碳单位的有机物都可以转变为琥珀酸,然后参加到TCA循环当中形成草酰乙酸,作为糖异生的原料,转变为葡萄糖或者其他糖类,也可以作为各种氨基酸合成的碳骨架。
那么乙醛酸循环的意义所在究竟在哪呢?我认为主要是直接打通乙酰CoA和琥珀酸的渠道,如果没有乙醛酸循环,虽然我们在TCA循环似乎也可以通过乙酰CoA变到琥珀酸,但是其实在循环转变的过程当中就已经经历了两次的脱羧反应,所以将琥珀酸脱离反应(或者经历后续反应生成的草酰乙酸脱离反应)就会使得TCA的中间体流量减少。所以乙醛酸循环就可以使得在不减少TCA流量的基础上,乙酰CoA可以转变为草酰乙酸,进而支持后续的糖异生或者其他反应。
所以乙醛酸循环不是一个目的在释放能量的过程,而是将二碳单位转变为四碳单位的合成路径。
对于植物来说,在富含油脂的种子发芽的时候,脂肪酸就可以转变为乙酰CoA,如果没有乙醛酸玄循环体,但是这里的乙酰CoA要么直接被TCA应用分解释放能量,要么储存(一般也不会),但是如果有乙醛酸循环体的作用的话,那么就可以促使分解得到的乙酰CoA先通过乙醛酸循环途径得到琥珀酸,然后接着经过TCA途径得到草酰乙酸,进一步就可以发生糖异生,从而合成葡萄糖,这样有助于提高种子细胞内的渗透压,从而促使其吸水萌发。
乙醛酸循环的调控
乙醛酸循环与TCA分叉的关键在于异柠檬酸,如果经过异柠檬酸裂合酶作用那么就进入了乙醛酸循环,如果是经过异柠檬酸脱氢酶的作用,那么就进入了TCA,至于进入哪一个循环受到磷酸酶和激酶的调控。一般来说,异柠檬酸裂合酶和异柠檬酸脱氢酶的活性是拮抗的,所以在细胞处于低能状态的时候,会抑制裂合酶的活性,抑制激酶活性激活磷酸酶活性从而激活脱氢酶,开启TCA,产生能量;如果处于高能状态,会激活激酶活性抑制磷酸酶活性从而抑制脱氢酶,激活裂合酶活性,从而开启乙醛酸循环。