Chapter1 转录网络
1.3 转录网络的因素
细胞是由数千种蛋白质组成的集成装置。每种蛋白质都是一个纳米级分子机器,能以极高精确度执行特定任务。细胞持续监测环境,并计算每种蛋白质所需数量,这一信息处理功能主要由转录网络执行
-
外部信号:物理参数(eg.温度),其他细胞的信号分子,有益的营养物质等 + 细胞内部状态信息
-
内部信号:内部状态信息
细胞通过产生合适的蛋白质响应这些信号,这些蛋白质作用于内外部。为表征这些环境状态,细胞采用一类称为转录因子的特殊蛋白质作为符号。转录因子可在活性态和非活性态之间转变
转录因子通过与受调控基因启动子中的特定位点结合来影响转录速率。结合后改变RNA聚合酶与启动子结合并产生mRNA分子的单位时间概率。既可以加快转录速率,也可以降低。
转录因子本身由基因编码,这些基因收到其他转录因子的调控,后者又可能被更上游的转录因子调控,以此类推,这一系列相互作用构成转录网络,该网络描述了细胞所有调控性转录相互作用

输入信号到达---->转录因子活性改变X变为X---->导致蛋白质生成速率改变---->其中一些是转录因子---->激活其他基因,以此类推*
网络中箭头的进化速度(X->Y:X产生的蛋白质是一种转录因子,可以与基因Y的启动子结合来控制基因Y的转录速率)远远快于基因编码区的进化速度。这类相互作用可以分为两种类型:
- 转录因子与启动子结合后提高转录速率---->激活
- 转录因子与启动子结合后降低转录速率---->抑制
锐角箭头代表激活(带"+"号),平角箭头代表抑制(带"-"号);箭头上不仅带符号,还承载着相互作用强度的数字。转录因子对靶基因的作用强度由输入函数描述:
rate of production of Y=f(X∗)rate\ of\ production\ of\ Y = f(X^*)rate of production of Y=f(X∗)
- X∗X^*X∗:激活态的转录因子浓度
- YYY:单位时间内蛋白质Y的分子数量
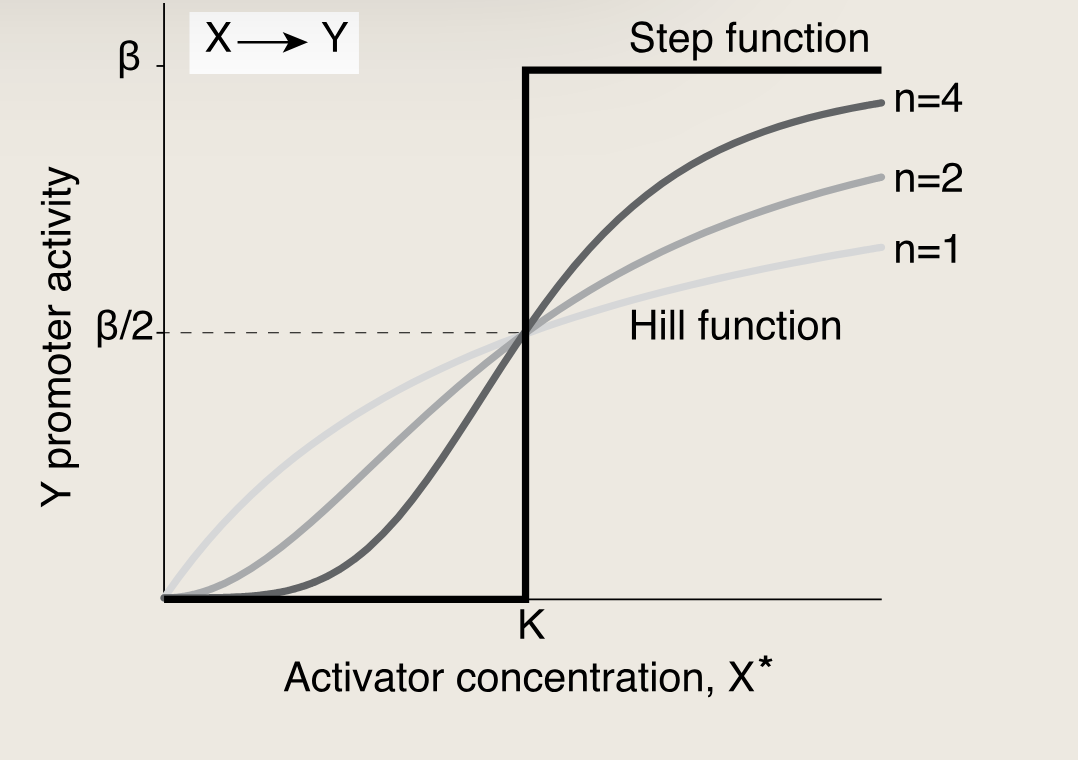
基因的转录调控可以用一个单调的函数来定量描述,Hill函数是其中一个典型代表:
KaTeX parse error: \tag works only in display equations
- KKK:激活系数,具有浓度量纲,当活性转录因子浓度达到K时,靶基因的表达到达半最大值(一半的最大转录速率)。K的值取决于转录因子与启动子结合位点的化学亲和力等因素
- β\betaβ:最大启动子活性,当X*浓度远高于K时候,表达达到最大值
- n:决定了输入函数的陡峭程度,n越大曲线越接近阶梯状
对于抑制因子(X------∣YX------|YX------∣Y),Hill函数是一个递减曲线。其形状取决于三个相似的参数:
KaTeX parse error: \tag works only in display equations
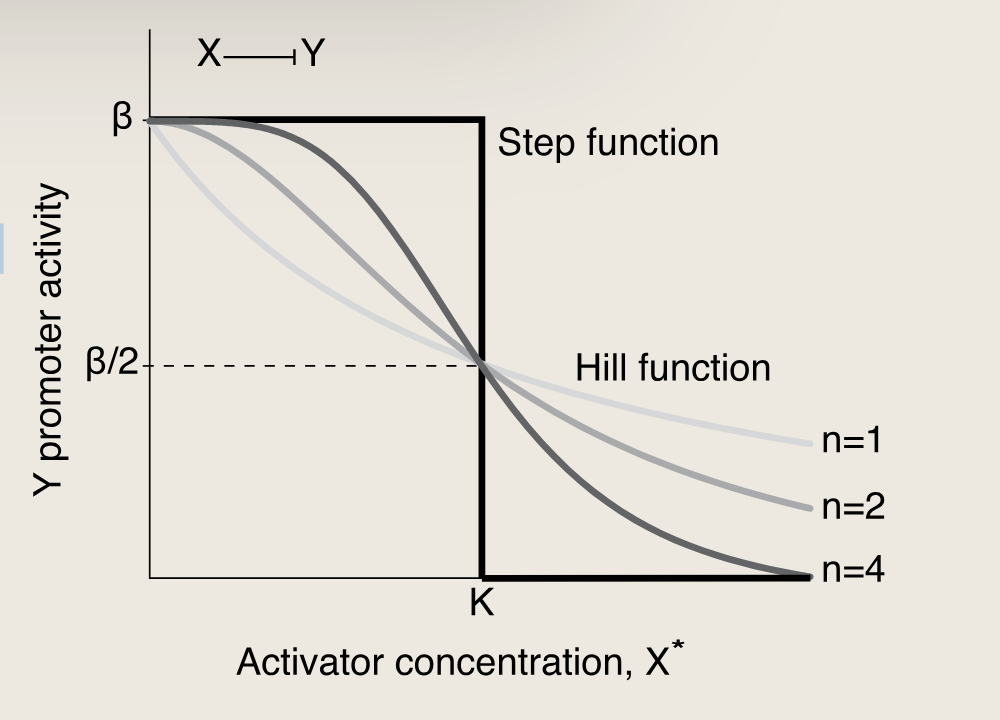
抑制因子不与启动子结合时才允许基因进行高效转录。
Hill函数用于详细建模,为了数学上的清晰性,有时使用更简单的函数来捕捉这些输入函数的本质行为是有益的。可以使用阶跃函数来描述,当X*超过阈值K时,该函数发生阶跃:
- KaTeX parse error: \tag works only in display equations
- θ\thetaθ等于0或者1,取决于括号中的逻辑陈述
- KaTeX parse error: \tag works only in display equations
使用逻辑输入函数,动态方程更容易求解
许多基因受到多个转录因子的调控,它们是网络中有两条或更多入边的节点。启动子活性是不同输入转录因子的多维输入函数
eg:考虑由两个激活因子调控的基因。许多基因都需要两个激活因子蛋白都结合到启动子,才能表现出高表达。这类似一个**"与门"**
- KaTeX parse error: \tag works only in display equations
对于其他基因,与其中一个结合就足够了,这类似一个**"或门"*
- KaTeX parse error: \tag works only in display equations
不是所有基因都有类似布尔的输入函数。eg:一些基因显示出一个SUM输入函数,其中输入是加法的:
- f(X+,Y+)=βxX++βyY+f(X^{+}, Y^{+}) = \beta_x X^{+} + \beta_y Y^{+}f(X+,Y+)=βxX++βyY+
1.4 简单调控的动态与响应时间
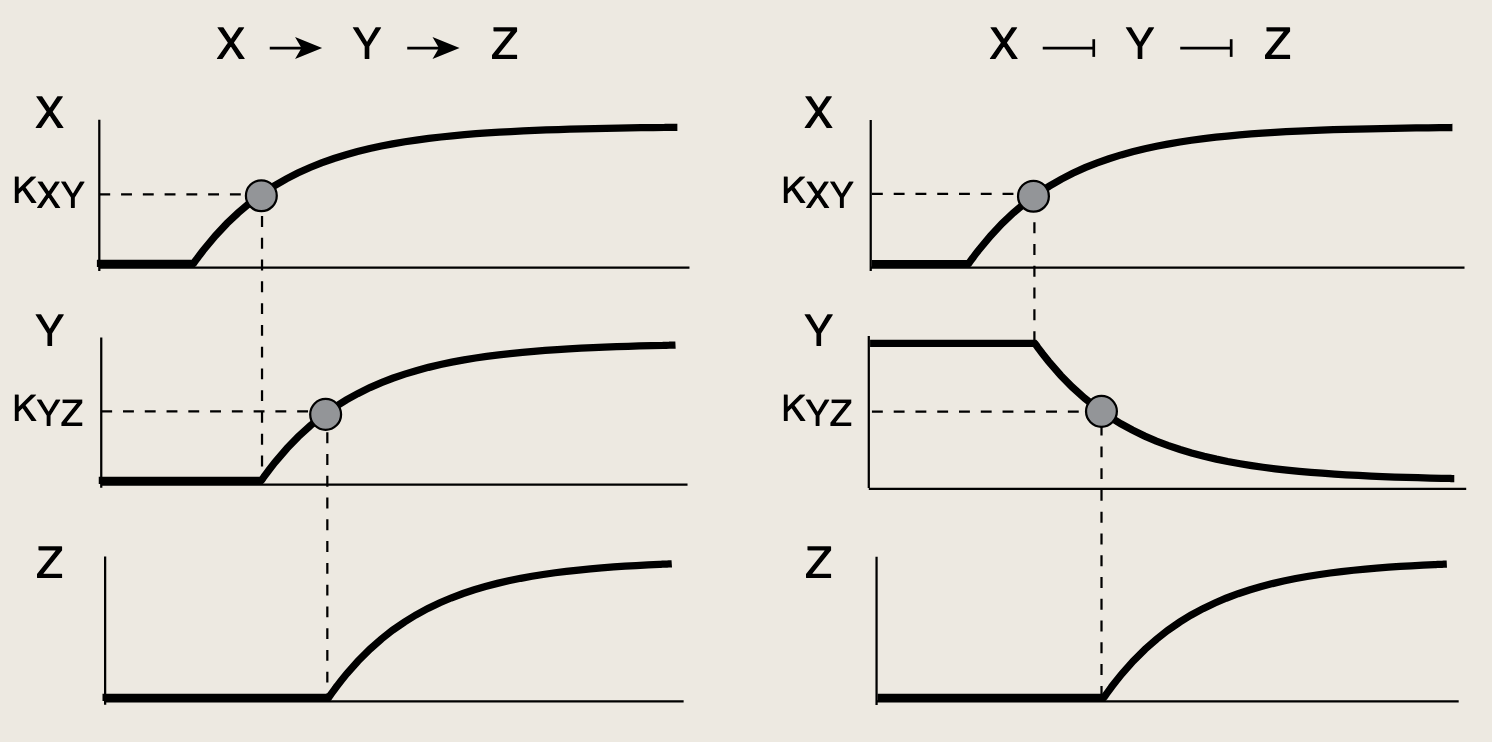
X ----> Y:转录因子X调控基因Y。一旦X被信号激活,Y的浓度便开始变化。计算蛋白质Y的浓度动态变化及其响应时间。
当信号Sx出现时,X迅速变为活性形式X,并结合Y的启动子,Y开始转录,mRNA被翻译,蛋白质Y开始积累*
- 在没有信号输入时,X也存在,但并非激活态,无法影响Y的转录
细胞以速率β\betaβ生产蛋白质Y,Y的生产受到两个过程的平衡:蛋白质降解(由细胞内特定蛋白将其特异性破坏,降解速度为αdeg\alpha_{deg}αdeg)和稀释(由于生长过程中细胞体积增加导致浓度降低, 稀释速率为αdil\alpha_{dil}αdil),随时间的总移除率:
- KaTeX parse error: \tag works only in display equations
Y的浓度变化源于其生产与清除之间的差值,由以下方程描述:
- KaTeX parse error: \tag works only in display equations
αY\alpha YαY代表浓度Y乘以每个蛋白质Y被移除的时间概率。在稳定态下,Y达到恒定浓度YstY_{st}Yst。通过求解导数为0点,即可得到稳态浓度是生产速率与移除速率之比:
- Yst=β/αY_{st} = \beta / \alphaYst=β/α
生产率β\betaβ越高,蛋白质稳态浓度越高;移除率α\alphaα越高,稳态浓度越低。
如果移除输入信号,那么β=0\beta = 0β=0,Y的生产停滞,方程在β=0\beta=0β=0时的解是Y浓度的指数衰减:
- KaTeX parse error: \tag works only in display equations
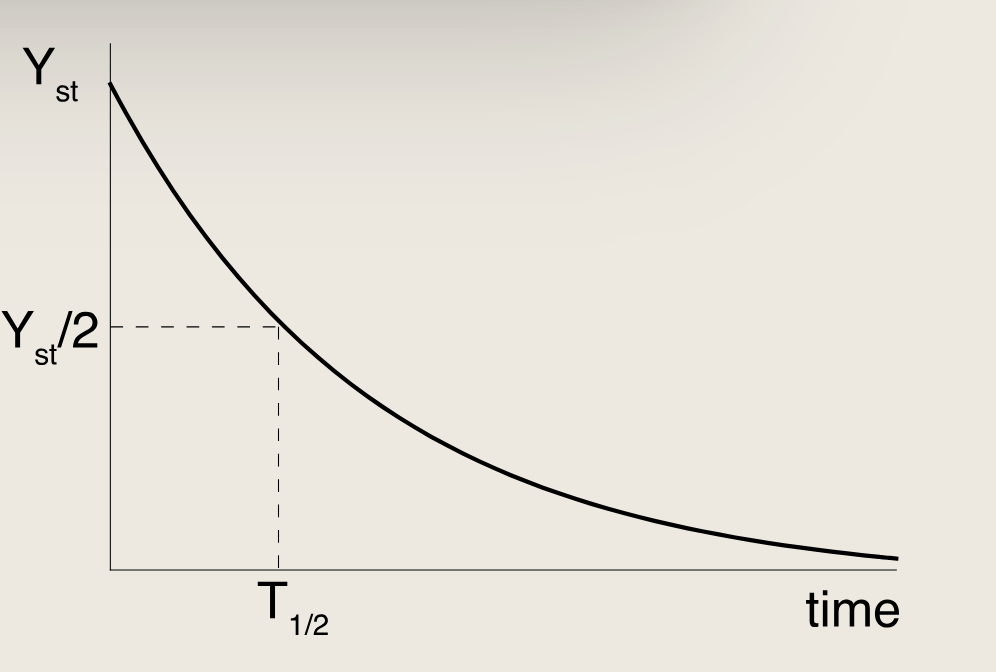
衡量Y的衰减速率的重要指标是响应时间(从YstY_{st}Yst下降到Yst/2Y_{st}/2Yst/2所需的时间),该时间与移除速率成反比:
-
-
T1/2=log(2)αT_{1/2} = \frac{\log(2)}{\alpha}T1/2=αlog(2)
去除速率α\alphaα直接决定响应时间,快速去除可使浓度迅速变化;生产速率β\betaβ只影响稳态水平,不影响响应时间
一个未受到刺激的细胞中Y=0,当施加信号后,蛋白质Y开始积累,如果一个未受刺激的基因突然被强信号Sx激活,动态方程将导致其趋近于稳态:
- Y(t)=Yst(1−e−αt)Y(t)=Yst(1−e^{−αt})Y(t)=Yst(1−e−αt)
许多蛋白质在生长细胞中不会被主动降解(αdeg=0α_{deg} = 0αdeg=0),这类蛋白质被称为稳定蛋白质。稳定蛋白质的生成通过生长细胞体积增大引起的稀释效应来平衡,即 α=αdilα = α_{dil}α=αdil。对于这类稳定蛋白质,其响应时间等于一个细胞世代时间。为理解这一点,设想一个细胞产生某种蛋白质后,突然停止生产(β = 0)。细胞继续生长,当其体积加倍时,分裂为两个子细胞。因此,经过一个细胞世代时间 τ 后,蛋白质浓度已降低 50%,故有:T1/2=log(2)/αdil=τT_{1/2} = log(2)/α_{dil} = τT1/2=log(2)/αdil=τ,即响应时间为一个细胞世代(1.4.9)。这是一个有趣的结果。细菌的细胞世代时间约为 30 分钟到数小时,而动植物细胞的世代时间通常为一天或更长。可以预见,那些用于响应营养物和胁迫等信号的转录网络,其反应速度应远快于细胞世代时间,否则只有细胞的子代才能从中获益。但正如我们所看到的,对于稳定蛋白质而言,响应时间恰好为一个细胞世代时间。因此,响应时间可能成为限制因素,对设计高效的基因电路构成约束。
Chapter2 自调控:一种网络模体
本章目标:
- 定义一种在复杂网络中检测构件模式(block-building)的方法,称为网络模体(network motif)
- 研究转录网络中最简单的网络模体,即负自调控
- 证明该模体具有实用功能:加速转录相互作用的响应时间,并增强其对噪声的稳定性
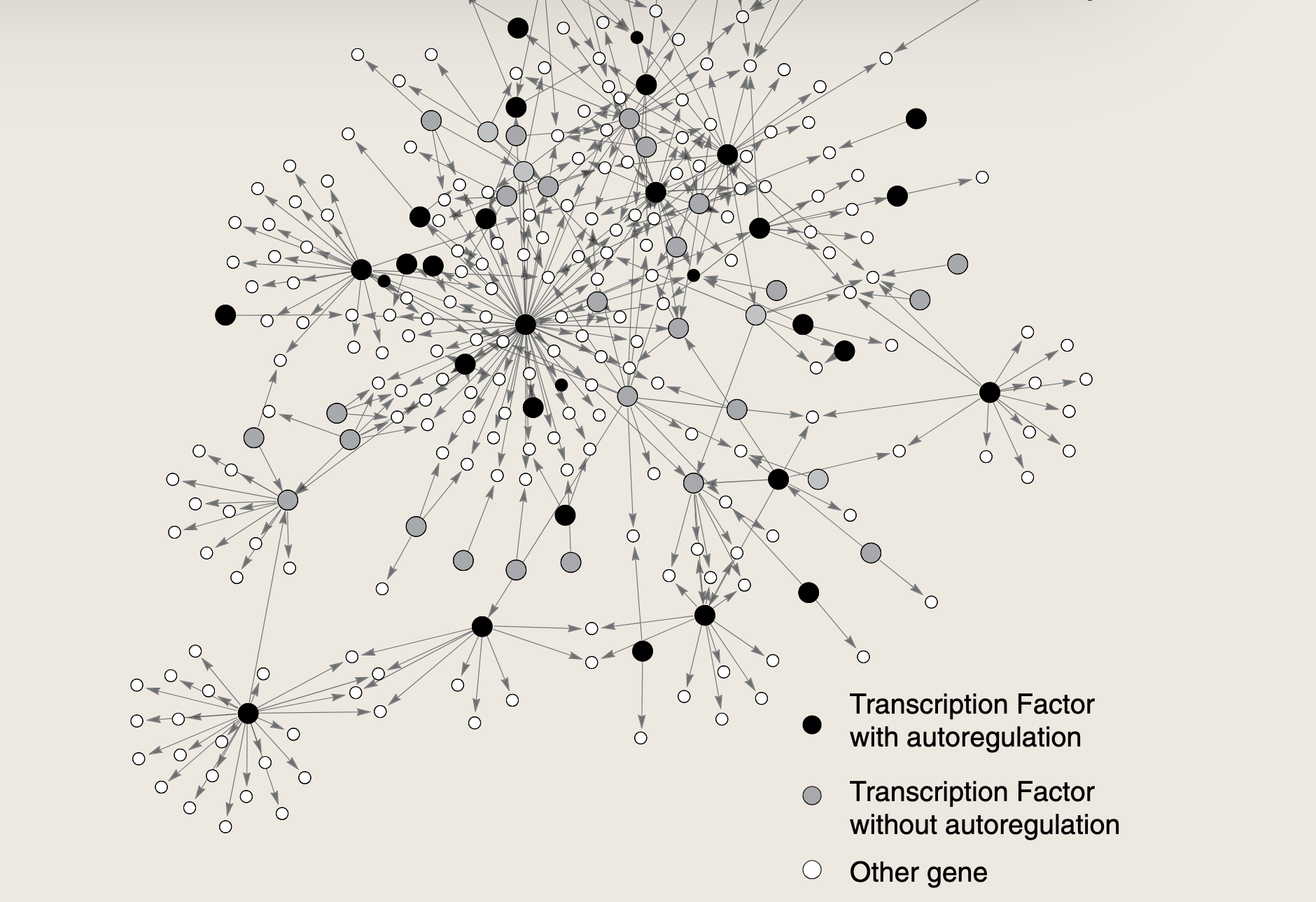 如上图所示大肠杆菌的部分转录网络
如上图所示大肠杆菌的部分转录网络
通过比较真实网络和与真实网络具有相同特征(eg:节点数与边数)的随机网络,捕捉有意义的模式
如果某种网络模体在真实网络中出现的频率远高于随机网络,它一定是因为其为生物体带来的某种优势而被选择保留的;否则它早已被淘汰。
自调节是一个网络模体
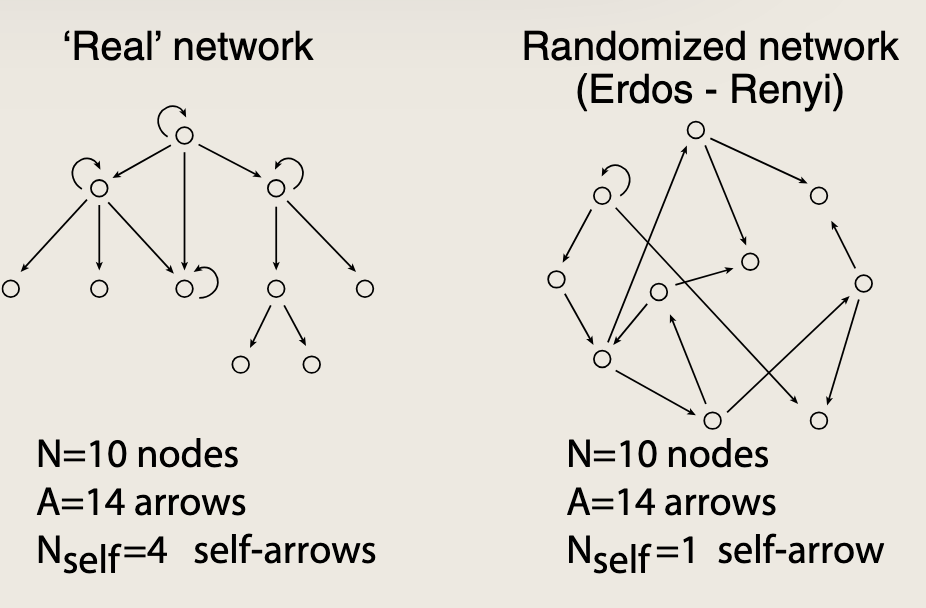
示例中的大肠杆菌网络包含40个自箭头,这些自箭头对应于调控自身基因转录的转录因子。基因由其自身基因产物调控的现象称为自生控制 ,或者称为自调控 。网络中的自调控蛋白中,有34个是抑制自身转录的抑制因子,即负自调控。
网络模体是指网络中出现的频率远高于随机水平的重复模式
该模体是否具有实用功能?通过比较负自调控基因与简单(非自)调控基因来探究。比较标准的系统响应时间 ,简单调控基因的响应时间由其取出速率α\alphaα决定:
- T1/2=log(2)αT_{1/2} = \frac{\log(2)}{\alpha}T1/2=αlog(2)
对于稳定蛋白质,其响应时间等于细胞世代时间
负自调控网络模体加速基因转录响应时间
当转录因子X抑制其自身的转录时,即发生负自调控。X与其自身启动子结合,从而抑制mRNA的生产。X的浓度越高,自身的生产速率越低。
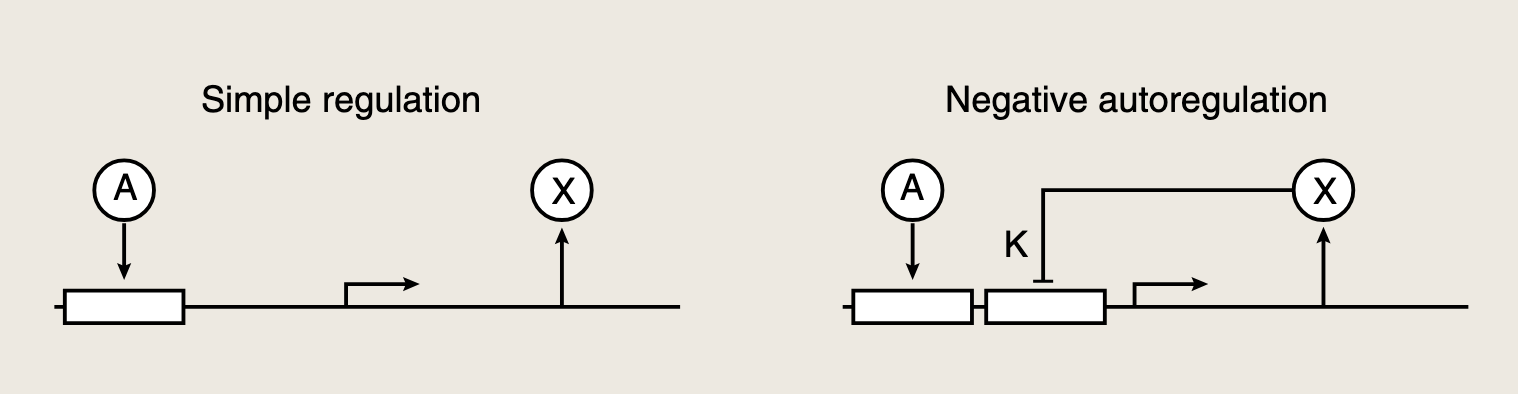
X的变化由其生产速率f(X)和移除速率α\alphaα描述:
- KaTeX parse error: \tag works only in display equations
f(X)是X的递减函数
- KaTeX parse error: \tag works only in display equations
当X远小于抑制系数K时,启动子处于自由状态,生产速率达到最大值β\betaβ;当抑制因子X浓度高时,转录不发生,即f(X)~0。
采用逻辑近似:
- X > K,则产量为0
- X < K, 则产量达到最大值
可使用阶跃函数θ\thetaθ来描述:
- f(X)=βθ (X<K)f(X) = \beta \theta\ (X < K)f(X)=βθ (X<K)
当X的水平达到自抑制阈值(X = K)时,生产便会停止,因为一旦X超过K,生产速率为0。若系统中存在延迟,则会在X=K附近产生小幅度震荡。延迟导致X略微超过K以上(X的浓度恰好到达阈值K时,已有的mRNA和正在翻译的核糖体仍然在继续生产新的X蛋白质,负反馈的抑制信号尚未完全生效,X的浓度会继续上升),随后生产停止,X水平下降直至低于K,此时生产再次启动,循环往复。对于实际的输入函数f(X),除非延迟非常长,这些震荡通常会被阻尼。也就是说X实际上会锁定在一个等于其自身启动子抑制系数的稳态水平:
- Xst=KX_{st} = KXst=K
刚开始XXX很小,移除部分可以忽略不计,所以在自抑制下,X一开始猛然增长;到达阈值K后,由于mRNA转录等过程已经进行,所以X会先上升,超过K;X停止生产,由于移除的存在,X会逐渐下降,低于K;此时又开始往复。

-
NAR代表自抑制的曲线,simple代表一般的曲线
-
βT1/2NAR=K/2βT_{1/2}^{NAR}=K/2βT1/2NAR=K/2
-
T1/2NAR=K/2βT_{1/2}^{NAR}=K/2\betaT1/2NAR=K/2β
-
K 越大,目标浓度越高,需要更久;
-
β\betaβ 越大,生产越猛,响应越快;
-
所以负自调控可以通过增大 β\betaβ 来加速响应
最大非抑制启动子活性β\betaβ越强,响应时间越短,因此负自调控可以利用强启动子实现快速生产,然后通过自抑制在期望的稳态水平左右停止生产。
将自调节与简单调节进行比较
两种电路达到相同稳态的条件(简单调控:Xst=βsimple/αsimpleX_{st} = \beta_{simple} / \alpha_{simple}Xst=βsimple/αsimple, 负调控:Xst=KX_{st} = KXst=K)
- K=βsimple/αsimpleK = \beta_{simple} / \alpha_{simple}K=βsimple/αsimple
简单调控的响应时间:
- T1/2simple=log(2)αsimpleT_{1/2}^{simple} = \frac{\log(2)}{\alpha_{simple}}T1/2simple=αsimplelog(2)
负自调控电路的响应时间:
- T1/2NAR=K2βT^{NAR}_{1/2} = \frac{K}{2 \beta}T1/2NAR=2βK
通过增大负自调控的β\betaβ,可以让二者的响应时间之比更小
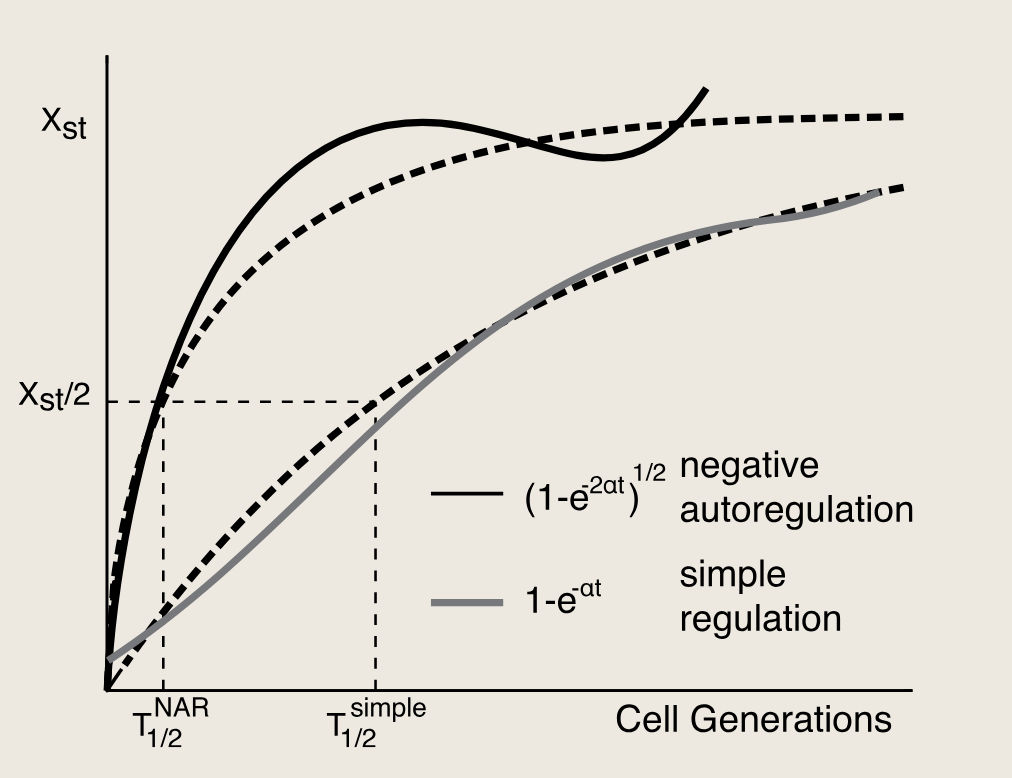
中负自调控设计的响应时间比简单调控快约七倍。加速的直观原因在于,负自调控利用强启动子实现快速初始上升,随后在期望的稳态水平K处自行停止。若采用相同强启动子的简单调控,则会达到过高的稳态水平,导致基因产物的不良过表达。因此,正如拥有强劲引擎和良好刹车才能成为快速驾驶者一样,负自调控电路相较于简单调控的加速响应已通过高分辨率基因表达测量得到实验证实。
首先考虑简单调控:
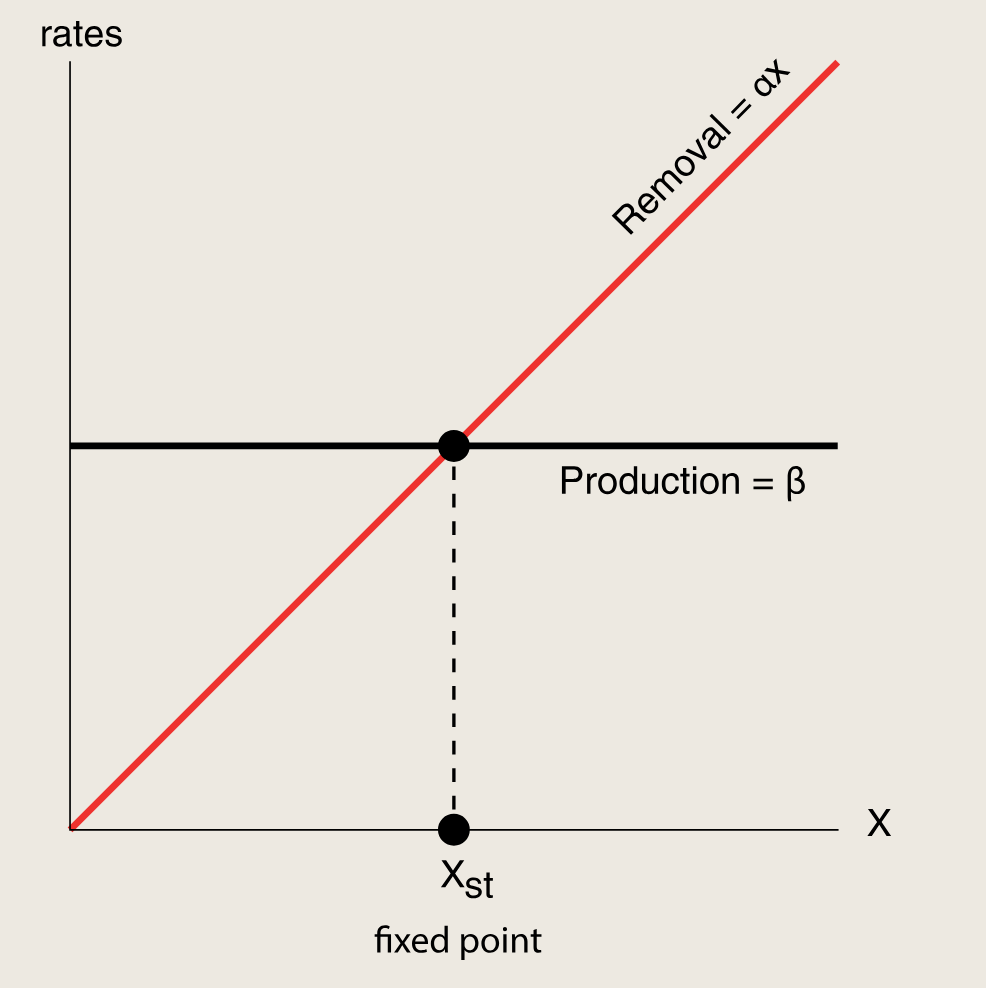
只要生产函数f(X)是随X增大而下降的,负自调控就会比简单调控更快接近稳态。在上图的相交点处,代表生产等于移除,此时X的浓度维持不变即达到稳态,这个不动点是全局稳定的:任何X值最终都会流回不动点。
对于响应时间 来说,X接近不动点的速度由两条曲线之间的距离决定,速度是时间导数:dXdt=生产−移除\frac {dX} {dt} = 生产 - 移除dtdX=生产−移除
接着考虑负自我调控:
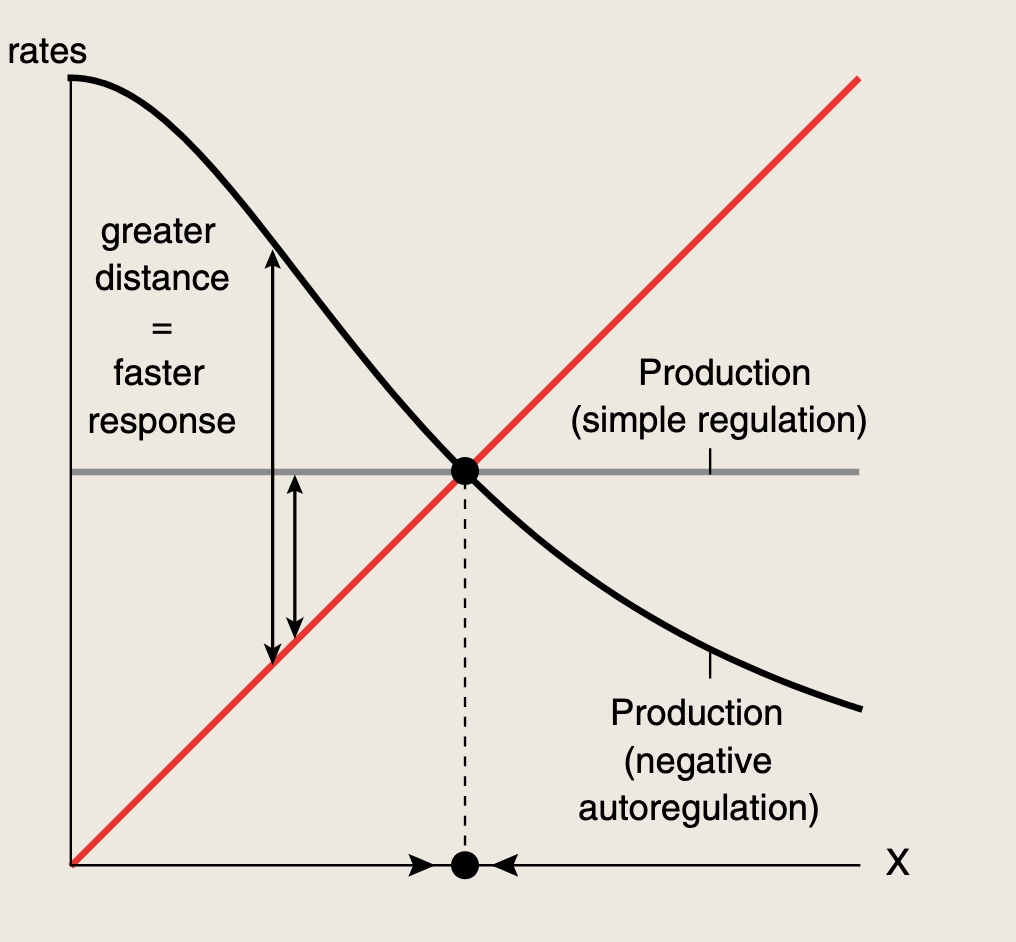
NAR中生产曲线与移除曲线之间的距离大于简单调控中的距离。因此,无论f(X)是什么形式,蛋白质水平X都将更快地移动到不动点,NAR加快了响应速度。
负自调控增加生产率波动的鲁棒性
负自调控对生产率的波动起到了缓冲作用
简单调控的稳态是:
- Xst=β/αX_{st} = \beta / \alphaXst=β/α
所以 beta 或 alpha 变一点,XstX_{st}Xst 就跟着变。细胞里 β\betaβ 会因为代谢状态、RNA 聚合酶数量、核糖体资源等发生波动,因此简单调控容易受到噪声影响。
负自调控在强抑制、陡峭输入函数下,稳态更接近:
- Xst=KX_{st} = KXst=K
K 主要由转录因子和 DNA 结合位点的亲和力决定,相对更"硬编码",细胞间波动较小。因此负自调控可以缓冲生产速率 beta 的变化,也能部分缓冲生长速率变化带来的 alpha 波动。
一句话总结:负自调控既能加速到达目标表达量,又能让目标表达量对生产/降解波动不那么敏感。
Chapter3 前馈循环网络模体
前馈环路的平均数量等于平均连接度λ\lambdaλ(每个节点的平均箭头数,λ=A/N\lambda=A/Nλ=A/N)的三次方,即λ3\lambda ^3λ3
- N:节点数
- A:箭头数
大肠杆菌转录网络中存在42个前馈环路,由3个节点组成的反馈环路为0;在具有相同平均连接度的对应随机化网络中,前馈环路的平均数量仅为约2个,反馈环路的平均数量小于1。
前馈网络是一种显著的网络模体,其出现频率远远高于随机水平;相比之下,三节点反馈环路则不是网络模体(在许多生物网络中它是一种反模体)。前馈网络是13种可能的的三节点模式中唯一显著的网络模体
在随机化力量的作用下,前馈环路为何仍被选择保留?它们是否执行了某种使生物体获得优势的功能?
前馈环路由转录因子X构成,X调控第二个转录因子Y,X和Y共同调控基因Z,因此,FFL包含两条平行的调控路径:从X到Z的直接路径以及经过Y的间接路径。FFL中的三条箭头可对应激活(+)或抑制(-),因此共有23=82^3=823=8中可能的FFL类型。
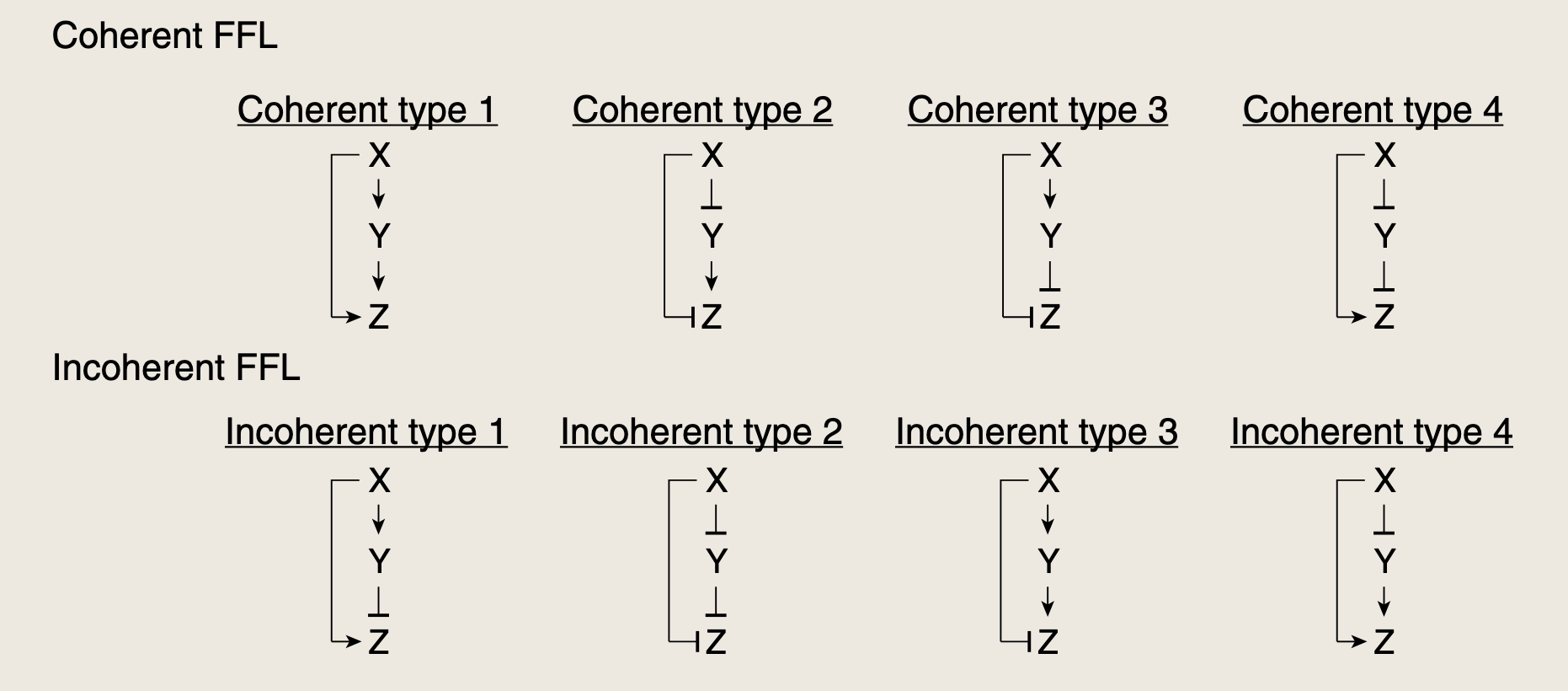
8种前馈环路可以分为两类:一致型 与非一致型。
- 在一致型中,间接路径的总体符号与直接路径相同。总体符号由路径上的箭头符号相乘得到。
- 一致型环路中包含偶数个负箭头
- 在非一致型中,间接路径的符号与直接路径的符号相反。
- 非一致型环路中包含奇数个负箭头
两种前馈型环路的出现频率远远高于其他6中,合计占所有前馈环路的80%以上。这两种类型分别是,一致型第一类前馈环路(C1-FFL),和非一致型第一类前馈环路(I1-FFL)
一致型
除了箭头的符号,要理解前馈环路的动力学,我们还必须知道两个调控因子X和Y在基因Z的启动子出如何整合输入,即基因Z的输入函数
我们将考虑两种在生物学上合理的逻辑函数:
- AND逻辑,即需要X和Y同时存在才能开启Z的表达
- OR逻辑,即X或Y中的任意一个就足以开启表达
FFL中的转录因子X和Y响应信号**SxS_xSx和SyS_ySy,在某些情况下,这些信号是直接与转录因子相结合的小分子;也有可能是先激活一条细胞内部的信号通路,然后这条通路再修饰X或Y。**
这些信号的作用时间尺度通常远快于FFL中转录相互作用的时间尺度
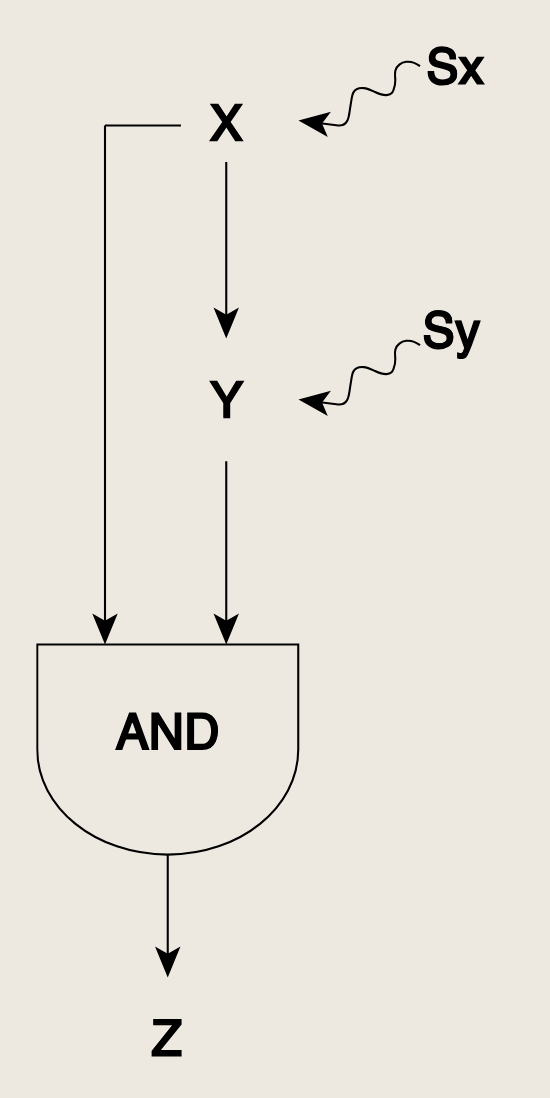
在此FFL中,所有三个相互作用均为正,对于Z启动子的输入函数,首先考虑AND逻辑,即需要两个激活因子X和Y都结合到Z的启动子上,才能启动蛋白质Z的生产。
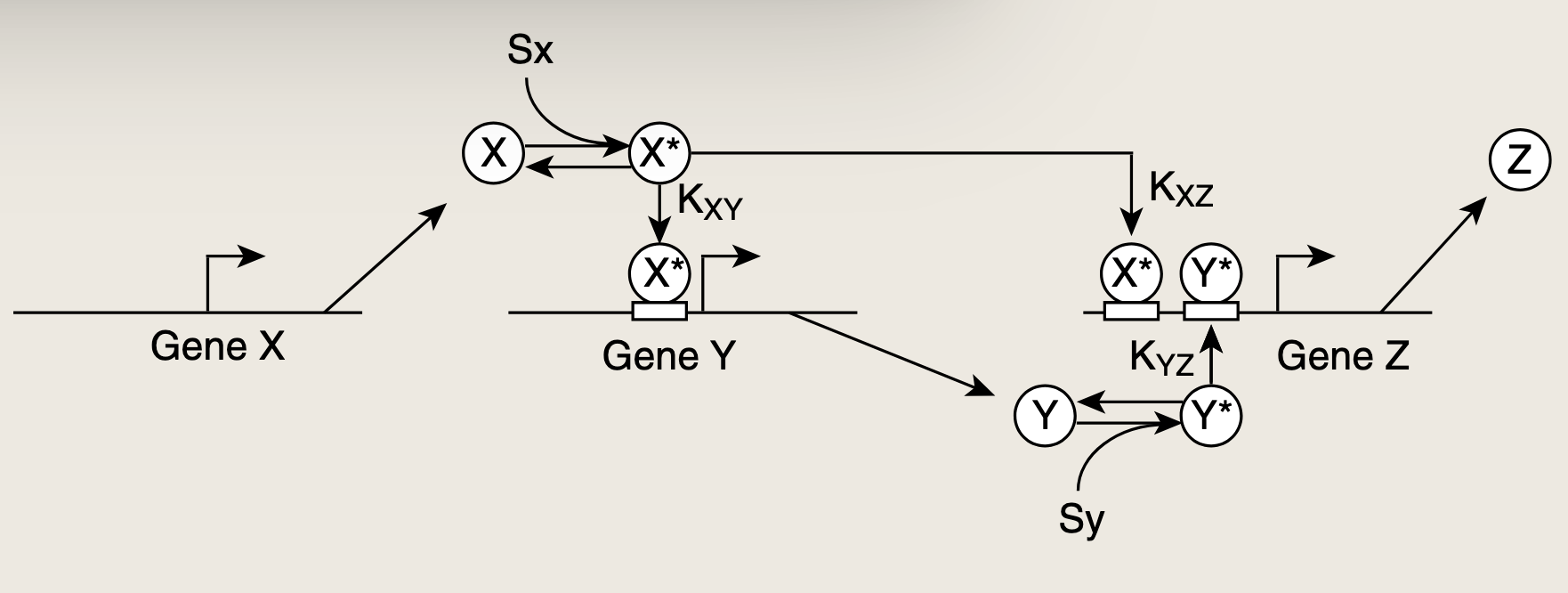
- 无信号时,X处于非活性形似。当信号Sx出现并触发X的激活,称为X的阶跃式刺激。XXX迅速转变为活性形式X∗X^*X∗。活性蛋白X与基因Y的启动子结合,启动蛋白质Y的生产。
- 与此同时其他的拷贝版的X∗X^*X∗与Z的启动子结合。但由于Z启动子的输入函数逻辑为AND,单独的X∗X^*X∗无法激活Z的生产。Z的生产需要X和Y同时结合。
- Z的激活还需要第二个输入信号SyS_ySy,从而使Y处于其活性形式Y∗Y^*Y∗。Y∗Y^*Y∗的浓度必须累积到足以超过基因Z激活阈值(KYZK_{YZ}KYZ)的水平。导致了蛋白质Z生产的延迟。
**对前馈环路的动力学进行描述,使用逻辑输入函数,这一关系使用阶跃函数θ\thetaθ**表示如下:
- production rate of Y=βyθ(X∗>KXY)\text{production rate of } Y = \beta_y \theta(X^* > K_{XY})production rate of Y=βyθ(X∗>KXY)
- KXYK_{XY}KXY:激活阈值
- βY\beta_{Y}βY:蛋白质Y产生速率
Y在X∗X^*X∗出现不久后便开始生产。Y的积累由之前已经熟悉的动态方程描述,该方程包含生产项和移除项:
- dYdt=βYθ(X∗>KXY)−αYY\frac{dY}{dt} = \beta_Y \theta(X^* > K_{XY}) - \alpha_Y YdtdY=βYθ(X∗>KXY)−αYY
Z的启动子由AND门输入函数住在。AND门可以被描述为一个两步函数的产物,因为两个调控因子都需要超过它们的阈值:
- KaTeX parse error: \tag works only in display equations
因此C1-FFL基因回路有三个激活阈值(箭头上的数字),Z的变化是带AND门输入函数的生产项与移除项的平衡:
- KaTeX parse error: \tag works only in display equations
C1-FFL是一种信号敏感型延迟元件
为分析C1-FFL的动力学,考虑对SXS_XSX的阶跃信号的响应:即SXS_XSX从无到有突然出现(ON阶跃),以及SXS_XSX突然消失(OFF阶跃)。为简化期间,我们始终假定信号SYS_YSY存在,因此转录因子Y处于其活性形式Y∗Y^*Y∗
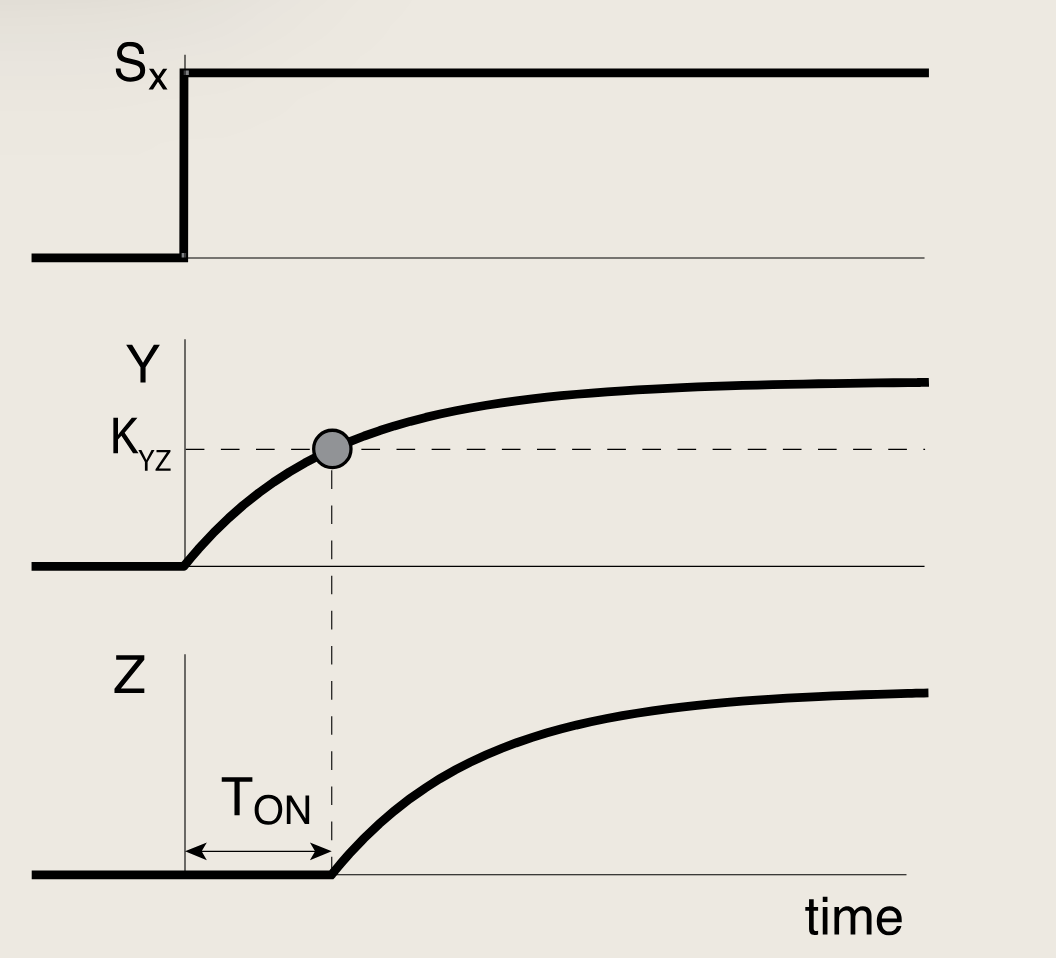
在SXS_XSX的ON阶跃后,Y开始以βY\beta_{Y}βY生成生成,Y的浓度开始指数收敛至其稳态水平Yst=βYαYY_{st}= \frac{\beta_{Y}}{\alpha_{Y}}Yst=αYβY:
- Y∗=Yst(1−e−αYt)Y^* = Y_{st}\left(1 - e^{-\alpha_Y t}\right)Y∗=Yst(1−e−αYt)
Z的蛋白生产受到一个与门函数控制,需要X,Y都超过阈值,所以Z在经历一段延迟后开始表达,该延迟TONT_{ON}TON即为Y达到起阈值所需的时间:
- Y∗(TON)=Yst(1−e−αYTON)=KYZY^*(T_{\mathrm{ON}})=Y_{st}\left(1 - e^{-\alpha_Y T_{\mathrm{ON}}}\right)=K_{YZ}Y∗(TON)=Yst(1−e−αYTON)=KYZ
可解出TONT_{ON}TON:
- TON=1αYlog(11−KYZ/Yst)T_{\mathrm{ON}}=\frac{1}{\alpha_Y}\log\left(\frac{1}{1 - K_{YZ}/Y_{st}}\right)TON=αY1log(1−KYZ/Yst1)
该方程描述了Z的延迟时间如何依赖于蛋白质Y的生化参数,包括蛋白质Y的去除速率,稳态水平与其激活阈值之间的比值。改变这些这些生化参数的突变可以对延迟时间进行调节。
当激活阈值KYZK_{YZ}KYZ超过Y的稳态水平时,TONT_{ON}TON趋近于发散,此时蛋白质Y永远无法达到激活Z的阈值 ,由于蛋白质生产速率的波动,YstY_{st}Yst容易受到细胞间差异的影响。
**因此,一个稳健的设计应该使阈值KYZK_{YZ}KYZ显著低于YstY_{st}Yst,以避免这些波动的影响。**在细菌中,典型参数所对应的延迟时间从几分钟到几小时不等。
SxS_xSx关闭阶跃之后无延迟,因为X失活并从Y和Z的启动子上解离。Z受到与门调控,只需一个输入关闭即可停止Z的表达,Z的生产随即停止,不存在延迟。
上述延迟表现称为信号敏感型延迟 ,其中信号敏感指的是延迟取决于阶跃符号的开启/关闭。信号敏感型延迟元件也可视为一种非对称滤波器。eg:若一个短暂出现的脉冲波SXS_XSX脉冲:
- 若开启脉冲的持续时间低于延迟时间,则C1-FFl不会产生Z的表达。因为在脉冲期间,Y蛋白没有足够的时间积累并跨过其激活阈值 ,只有大于TONT_{ON}TON的持久脉冲才能触发Z的表达。因此,此类FFL对开启脉冲启到持久性检测器的作用
- 另一方面,对于关闭脉冲能够立即响应。
- 与FFL相比,简单调控(不含FFL)无法滤除短输入脉冲,只要输出脉冲存在,Z的表达便会持续
细胞的环境常常波动,有时刺激可能以短暂脉冲的形式出现,而不应引发响应。C1-FFL可以提供一种过滤功能,在此类波动中具有优势。
与AND门C1-FFL不同的是,或(OR)门C1-FFL拥有对关闭阶跃的信号敏感型延迟
- 当SxS_xSx施加开启阶跃时,Z会立即被激活,因为只需要一个输入即可触发表达
- 相反,在关闭阶跃阶段会延迟失活,对于或门而言,需要两个输入都关闭才能失活,SxS_xSx关闭阶跃后,Y∗Y^*Y∗的衰减需要时间。因此,采用或门的C1-FFL也是一种信号敏感型延迟元件,只不过与AND门相反,即使输入信号短暂丢失,也可以维持Z的表达
非一致型
我们可以采用同一种方法理解另一种常见的前馈环路类型------非一致型第一类前馈环路(I1-FFL)。在该回路中,X激活基因Z,同时也激活Y,而Y则抑制Z。现考虑Y强烈抑制Z的情况:
-
在输入信号SxS_xSx施加一个开启阶跃后,X与Y和Z的启动子结合。由此Z的浓度开始上升,与此同时,抑制因子Y也在积累,当其浓度跨越其抑制阈值KYZK_{YZ}KYZ的时候,Y将关闭Z的生产,Z的浓度水平开始下降。
-
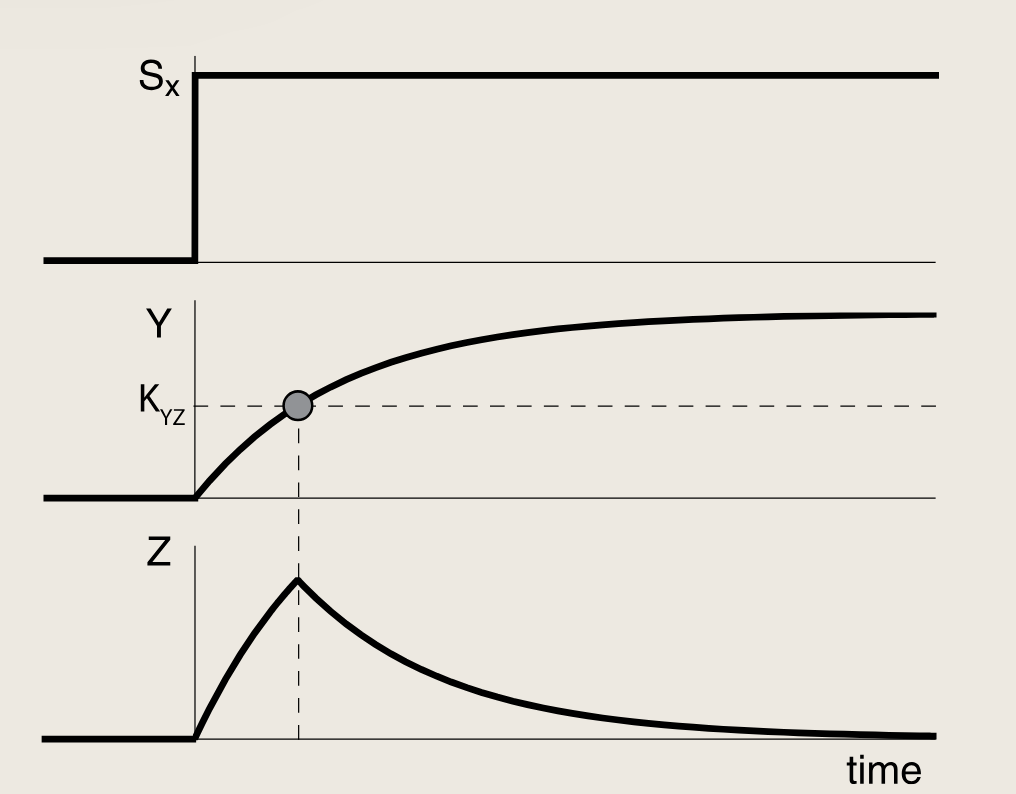
-
Y强烈抑制Z时,会产生一个脉冲(Z先升高,又被降回0)
-
Y部分抑制Z时,Z会收敛到一个稳态Zst,该稳态由X的激活作用与Y的抑制作用之间的相对强度共同决定
- 在此种情况下,可将I1-FFL与达到相同Zst的简单调控进行比较
- I1-FFL相比简单调控能够加快响应速度,因为I1-FFL允许快速上升(Y的浓度尚低的时候),随后由Y将其抑制在ZstZ_{st}Zst水平。这一原理与我们在负自调控中看到的强大引擎和强力刹车类似。
通过以上内容我们可以总结出三种加速基因表达响应时间的方法,通常从开启到生产足够蛋白质需要较长时间,三种方法是:
-
增加降解速率:让蛋白质快速降解,这样浓度变化更快,但代价是要多生产以维持稳态,形成"无用循环"。
-
负自调控:基因产物抑制自身表达,可以用强启动子快速生产,达到目标浓度后自行关闭,加速响应。
-
非一致型前馈环路(I1-FFL):通过一个延迟的抑制子来关闭快速生产,同样能加速开启响应,且适用于任何蛋白质(不限于转录因子)。
这三种方式可以相互结合,进一步提升速度。
非一致型第一类前馈环路可产生双相稳态响应曲线
- 简单调控中X->Z产生单调响应函数,输出ZstZstZst随着SxS_xSx上升而增加
- I1-FFL可以产生非单调响应:
- ZstZstZst先随着SxS_xSx上升,达到较高水平后转而下降,形成一条倒U形响应曲线。由于响应曲线包括上升和下降两个相位,因此称为双相响应。
- 当X激活Z所需的阈值低于激活Y的阈值时,I1-FFL中便会出现双相响应
- 在SxS_xSx较低时,阻抑蛋白Y未被激活,仅仅有X->Z这一支路在作用,响应曲线呈上升趋势
- 在信号强度较高时,Y被合成,ZstZ_{st}Zst随信号增强而下降
通过本章讨论的方法,可以分析其余六种前馈环路(FFL)各自的功能。分析表明,所有四种一致型前馈环路均可作为信号敏感型延迟元件,在输入信号的开启或关闭阶跃后产生延迟。所有四种非一致型前馈环路均可产生脉冲和双相响应,但具体细节有所不同。例如,非一致型第三类前馈环路可呈现一种先随输入信号下降、再上升的双相响应,形成U形而非倒U形的响应曲线。由于所有FFL都具有潜在有用的功能,因此为何C1-FFL和I1-FFL这两种类型出现频率远高于其他六种。
在13种三基因模式种,前馈环路(FFL)是大肠杆菌和酵母的转录网络以及高等生物感觉转录网络中唯一的一种网络模体。在FFL的8种可能组合中,最常见的是C1-FFL和I1-FFL。前者可以充当持久性检测器,消除短暂的输入脉冲;后者可在时间上和输入剂量上产生脉冲,并加速响应速度(该种加速作用可与其他加速机制协同使用),进化在不同基因系统和不同生物体中反复趋同于FFL。
Chapter4 时序程序和转录网络的全局结构
在之前的内容中,我们详细了解了其中两种模体:负自调控 和前馈回路。整个转录网络,仅由四类网络模体家族构成,这四类模体家族几乎解释了网络中所有的相互作用,将这些网络模体组合在一起,构建出转录网络的全局结构。
单输入网络模体(SIM)
在SIM网络模体中,一个主转录因子X控制一组靶基因Z1Z_1Z1, Z2Z_2Z2...ZnZ_nZn。SIM中的每个靶基因仅受一个输入调控 ;没有其他转录因子,调控其中任何一个基因。此外,SIM中所有基因的调控(激活/抑制)均相同 。SIM的最后一个特征是主转录因子X通常具有自调控特性。
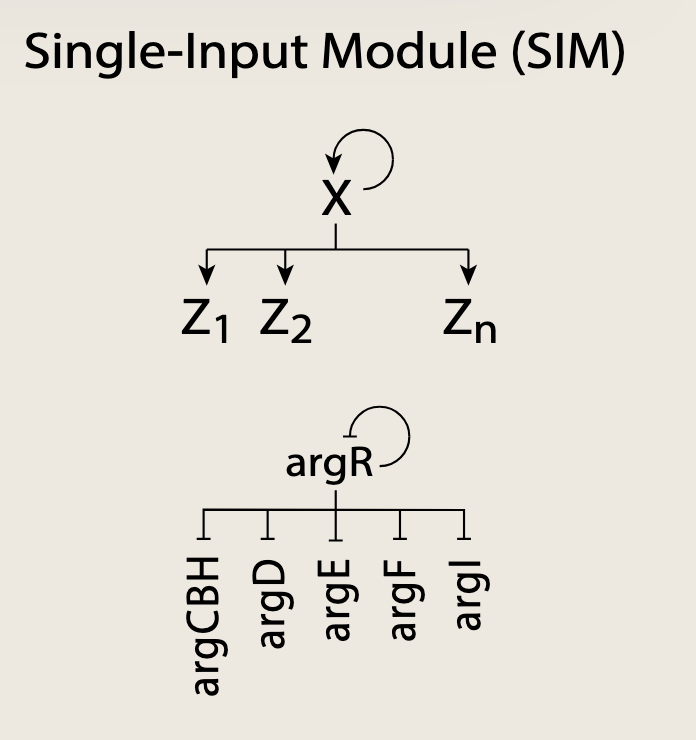
SIM最重要的作用是根据主转录因子感知到的信号来控制一组基因。SIM中的基因通常具有共同的生物学功能。
eg:SIM调控参与特定代谢途径的基因。这些基因编码的蛋白质以分子装配线的方式,逐步组装出一个所需的分子。代谢途径通常还表现出额外的反馈控制,这种控制以快速的时间尺度运动。该途径的最终产物往往是顶端转录因子的输入信号。最终产物还通常直接结合来抑制途径中的第一个酶,这种调节模式称为反馈机制
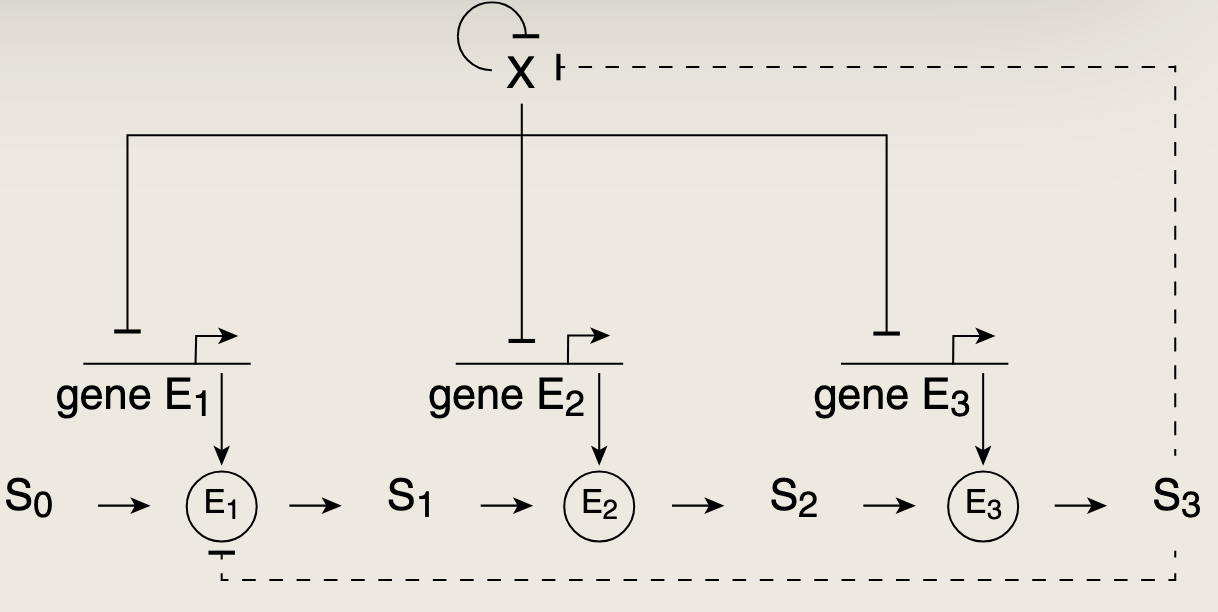
SIM能够产生时间性的表达,使基因按特定顺序依次激活。这种时间顺序基于靶基因ZiZ_iZi对转录因子X的不同阈值。当X的活性随时间逐渐变化时,会在不同时刻依次越过这些阈值KiK_iKi。X首先激活最低阈值的基因,紧接着激活次低,以此类推。X活性变化越快,它越过不同阈值的速度就越快,各基因激活之间的延迟也就越短。当X活性下降时,失活的顺序正好相反。SIM模块产生的时间顺序可以通过改变基因阈值的突变来调整
在这些全局定时响应中,基因通常由一个主调控因子以及负责更小子系统的额外辅助调控因子共同调控。即便网络模式并为严格的单输入模块。只要在关注的时间区间内除一个调控因此外其余所有调控因子的活性保持恒定,时间顺序仍可能通过主协调调控因子的作用而产生。
简而言之,SIM能够生成按需准时、具有后进先出顺序的时间程序:基因激活顺序与失活顺序相反。然而在许多情况下,激活顺序和失活顺序相同似乎更为理想:即最先开始启动的启动子也最先关闭,对于要求部件以特定顺序参与的组装过程而言,FIFO顺序更为可取。在该过程下,让早起基因在晚期基因之间关闭更为合理,以避免在晚期基因关闭后仍不必要地生产早期蛋白造成浪费。该种FIFO顺序可以通过多输出前馈环路实现。
双输出前馈环路与双扇结构
转录网络中发现的更大子图均可由最多包含四个节点的模体构建而成。可能的四节点模体共有199种,但只有其中的2种属于显著模体。这两种显著网络模体分别是:双输出前馈环 以及双扇结构的重叠调控模式。
双输出前馈环
- 在双输出前馈环中,X调控Y,且二者共同调控输出基因Z1Z_1Z1和Z2Z_2Z2。
- 扩展到更大子图时,这种结构可以泛化为多输出前馈环模式 :
- 由两级调控因子构成的级联结构共同调控一组输出基因
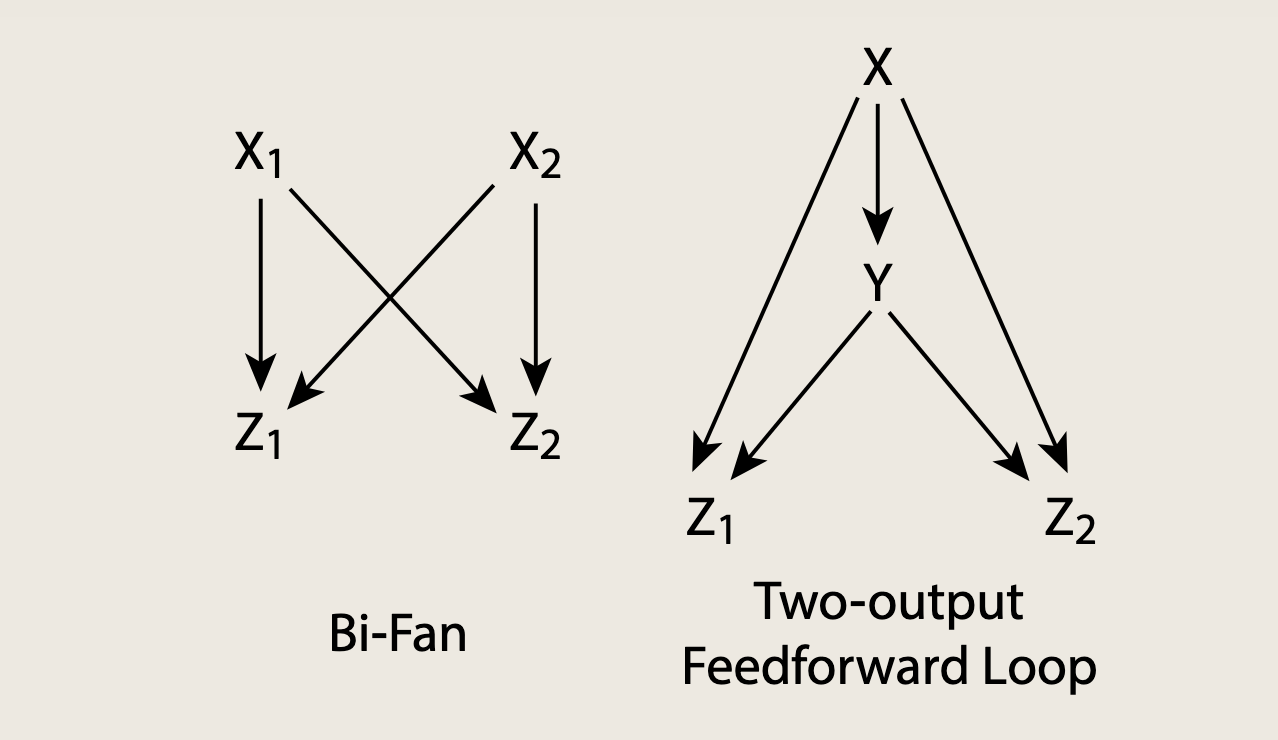
- 上图分别为双扇和双输出前馈环
多输出前馈环可生成先进先出(FIFO)时序程序,这与单输入模体所生成的后进后出顺序形成鲜明对比
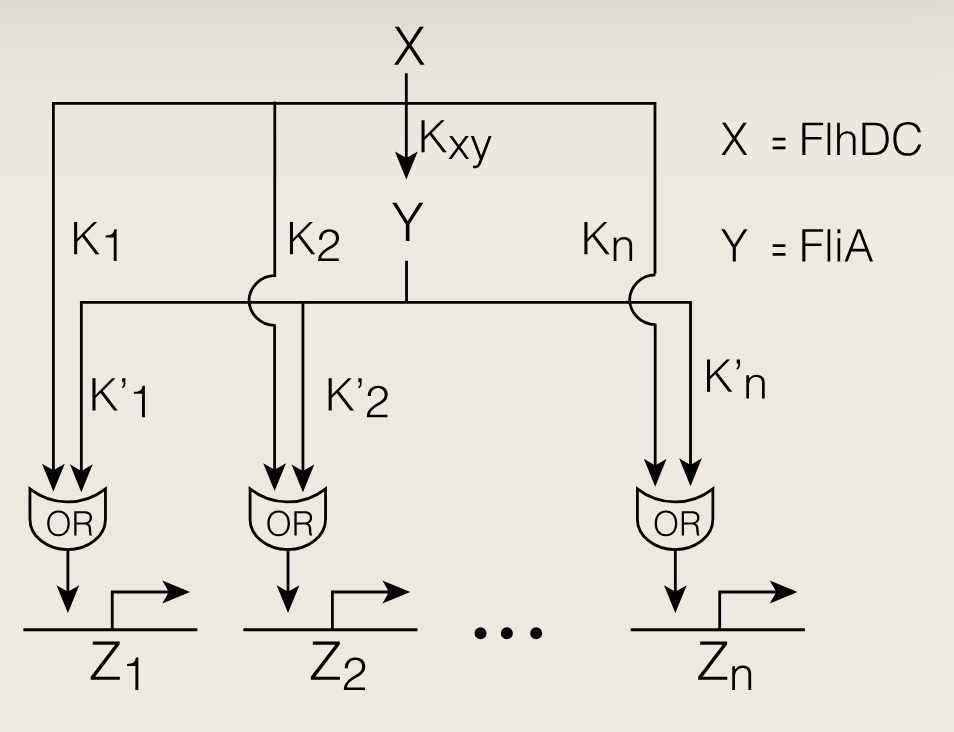
上图中,鞭毛马达操纵子受两个转录激活因子X和Y调控。主调控因子X激活Y,两者共同激活六个操纵子Z1Z_1Z1、Z2Z_2Z2、...、Z6Z_6Z6。在该多输出前馈环中,每个操纵子可在没有Y时被X激活,也可以在没有X时被Y激活。因此其输入函数类似或门。
若Y的阈值顺序与X的阈值顺序相反,则能实现FIFO顺序。即X的阈值满足**K_1 \< KaTeX parse error: Can't use function '' in math mode at position 4: K_2̲\*\*,Y的阈值应逆向排列\*\*...K_2'$**。
信号关闭时,X快速下降,但是Y下降较慢
多输出FFL还传递了所有前馈环的功能。每个输出节点都受益于FFL的符号敏感滤波特性
- eg:FFL延缓了X活性丧失后Z基因的失活,因此它会过滤掉X的短暂OFF脉冲,只有当X活动持续消失一段时间后才允许失效。
双扇结构
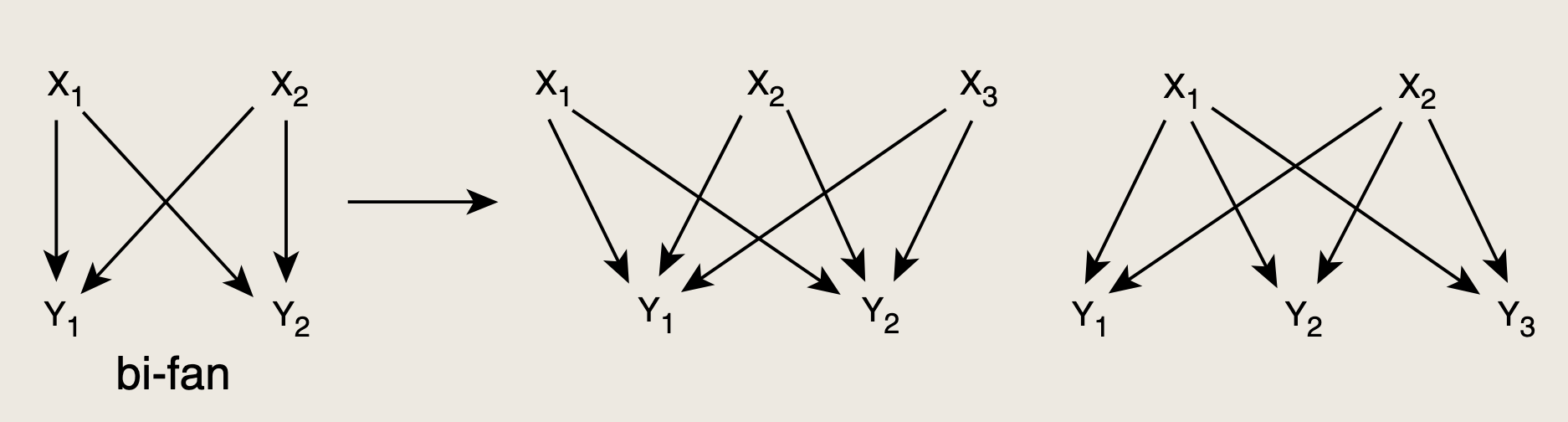
输入层与输出层间有多种多样连接。
这类模式是感觉网络中最后一种模体,称为密集重叠调控子(DOR),其中"调控子"指的是由特定转录因子调控的一组基因。
DOR由一排输入转录因子构成,以高度重叠的方式调控一排输出基因。DOR通常并非完全联通的,即并非每个输入都调控输出。但其连接密度远高于随机化网络中的模式。
可将DOR视为一种组合决策装置。它由一系列门控阵列组成,通过整合多个输入来计算每个输出基因的调控方式。要理解DOR的功能,需要掌握在各基因启动子处整合输入信号的多维输入函数知识。---->这构成了系统生物学领域尚未解决的挑战之一
通过将各类模体网络替换为不同的几何图形,我们可以得到粗粒化的转录网络,由该网络我们可以得出,大肠杆菌和酵母等生物的感觉转录网络均由单层DOR构成------这些DOR不形成级联,没有DOR出现在另一DOR的输出端,因此,网络完成的大部分计算发生在DOR内部的启动子层。
大多数的其他模体都被整合在DOR之中。前馈环和单输入模体均内嵌于DOR内,负自调控常与前馈环整合,并点缀在SIM主调控因子的周围。
迄今为止,所有的感觉转录网络,几乎所有基因都涵盖在这四类网络模体之下
下一章我们将讨论发育转录网络中出现的其他网络模体,这些网络引导着单细胞卵向着多细胞生物的转化过程
互锁前馈环
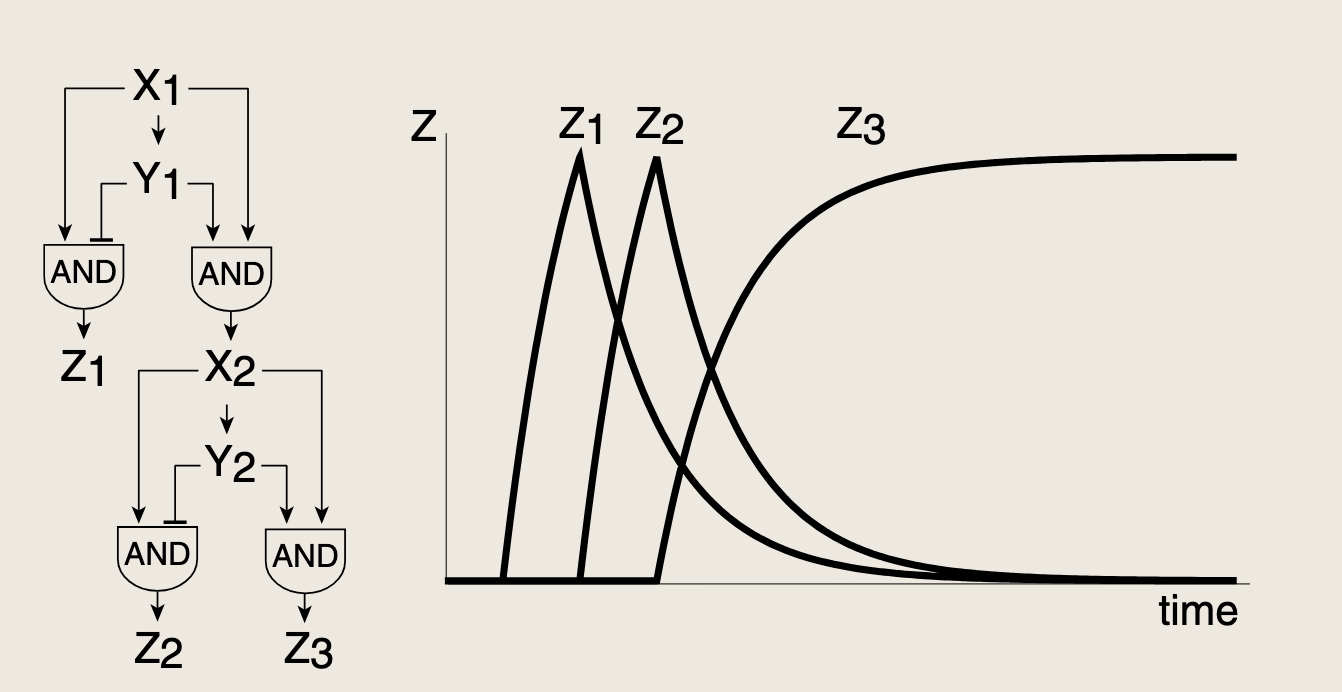
- X1X_1X1首先在饥饿信号下被激活,该转录因子与阻遏蛋白Y1Y_1Y1共同作用于第一类非一致型前馈环,控制一组输出基因Z1Z_1Z1;与此同时共同形成第二个前馈环,二者均为激活X2X_2X2所必须,从而构成一个具有与门逻辑的第一类一致性前馈环。该C1-FFL的存在使得只有当饥饿信号持续存在的时候,X2X_2X2才会被激活。随后,X2X_2X2在另一个I1-FFL中发挥作用:它既激活Z2Z_2Z2,也抑制其阻遏蛋白Y2Y_2Y2。这导致Z2Z_2Z2以相对于第一个脉冲延迟的时间产生一个脉冲。最后Y2Y_2Y2和X2X_2X2共同通过一个C1-FFL激活基因Z3Z_3Z3。
- 形成一个三波模式
**综上所述,不同生物的感觉转录网络由四类模体家族构成:自调控(autoregulation)、前馈环(FFL)、单输入模体(SIM)和密集重叠调控子(DOR)。**几乎所有的基因都参与这些模体。
Chapter5 正反馈,双稳态与记忆
5.1 发育网络中的网络模体
到目前为止我们关注的是为感知和响应外界刺激而构建的转录网络,这类转录网络存在于几乎所有细胞中。多细胞生物还拥有另一种转录网络,称为发育转录网络。这些网络调控细胞在转化为另一种类型时所发生的近乎不可逆的变化。
在若干中不同的生物体中,发育转录网络由若干强网络模体构成,它们展现出我们在感觉网络中发现的大多数模体。例入:FFL就是一种强网络模体。与感觉网络类似,发育网络中最常见的FFL类型是C1-FFL和I1-FFL。发育网络还呈现出显著的自调控模体和单输入模体。
除了这些模体,发育网络还展现出一些感觉转录网络中不常见的额外模体。
5.1.1 正自调控减缓响应速度并可导致双稳态
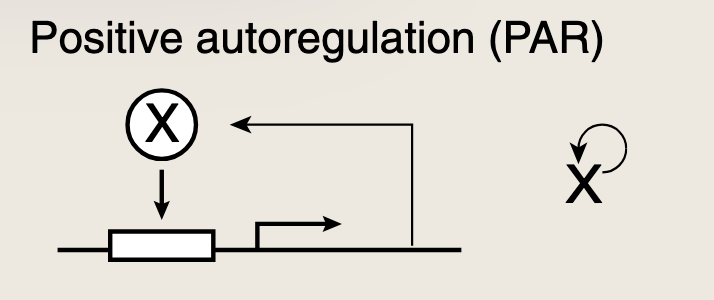
发育网络中**正自调控(PAR)**环的数量远远多于感觉网络中的。在正自调控中,蛋白质激活自身的转录,与负自调控相反,正自调控会减缓响应的时间:
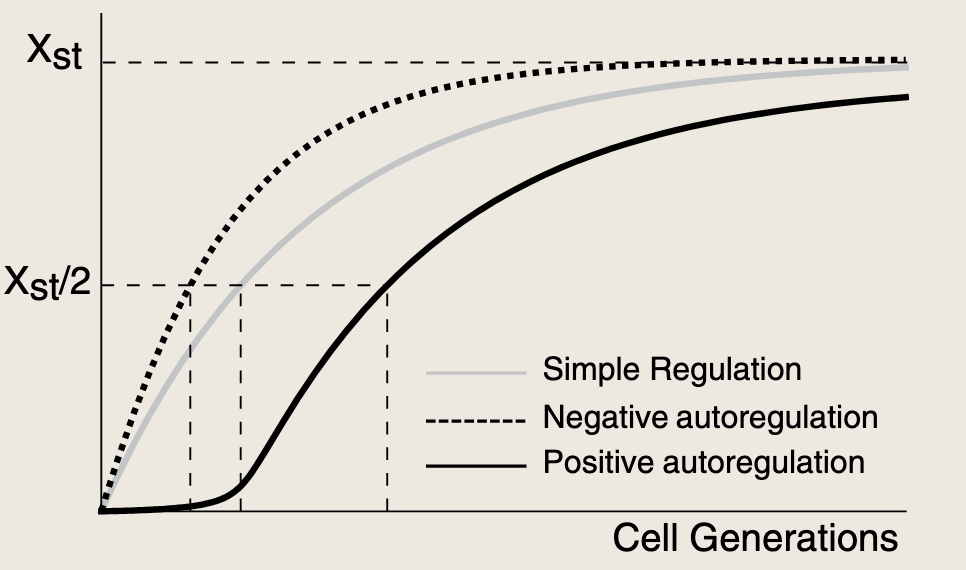
- 动力学过程最初缓慢,随着X水平的积累,它加速自身的生成,达到稳态一半的时间比简单调控更迟。
将正自调控过程中f(X)f(X)f(X)随X的浓度的变化与简单调控的相比较:
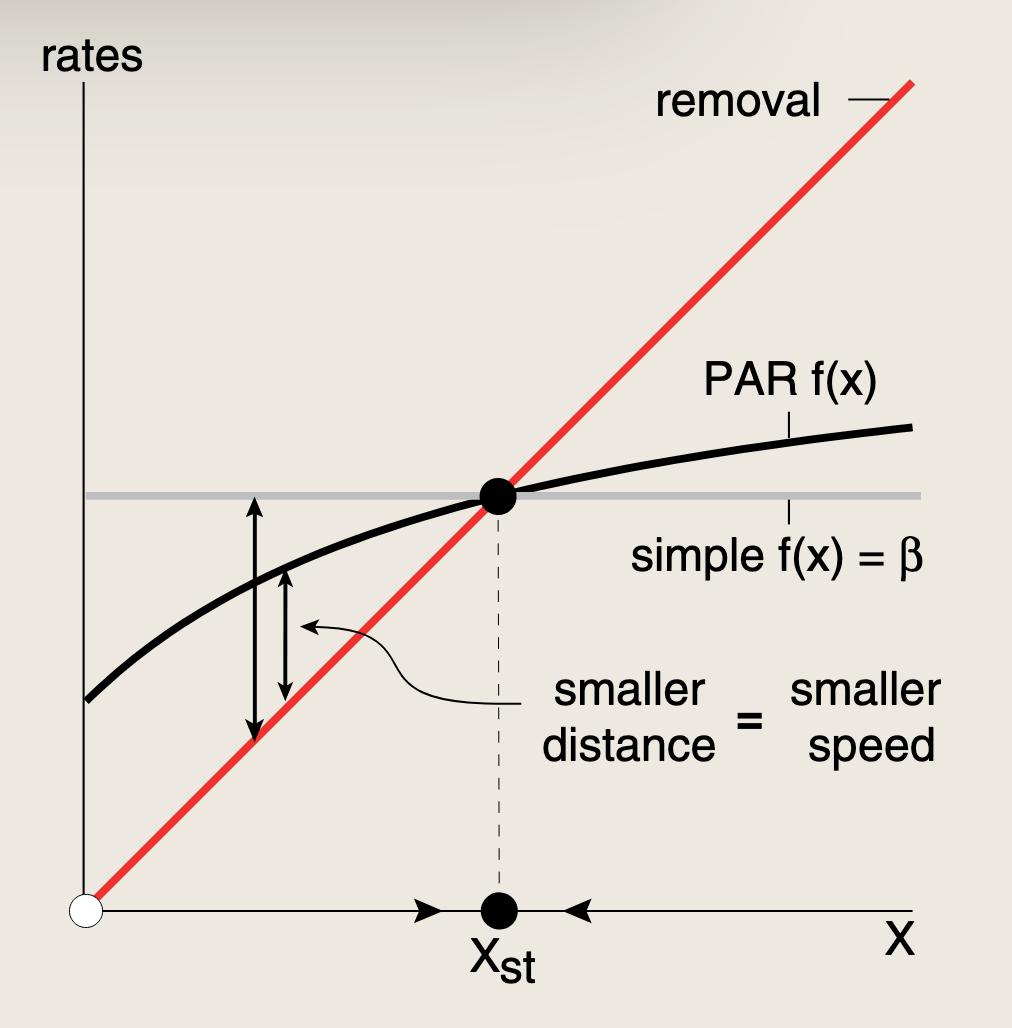
- 激活能力是有上限的
正自调控所提供的缓慢动力学在需要较长时间的多阶段过程中非常有用,例如发育过程。这些过程受益于不同阶段蛋白质生成之间的长时间延迟。缓慢的响应时间还有助于滤除输入信号中快速变化的噪声,慢响应能够整合这些噪声,使之相互抵消。
正自调控还能实现一项非凡功能:**它能够在两种状态之间做出清晰的决策,并将该决策长时间记忆下来。当自调控函数f(X)具有协同性(即S形曲线)时,PAR即可提供这种记忆。**若位置恰当,输入函数可以与移除曲线相交三次。(X 会激活自己的生产。如果这种正反馈足够强,生产函数f(X)f(X)f(X) 会变成比较陡的 S 形曲线。)
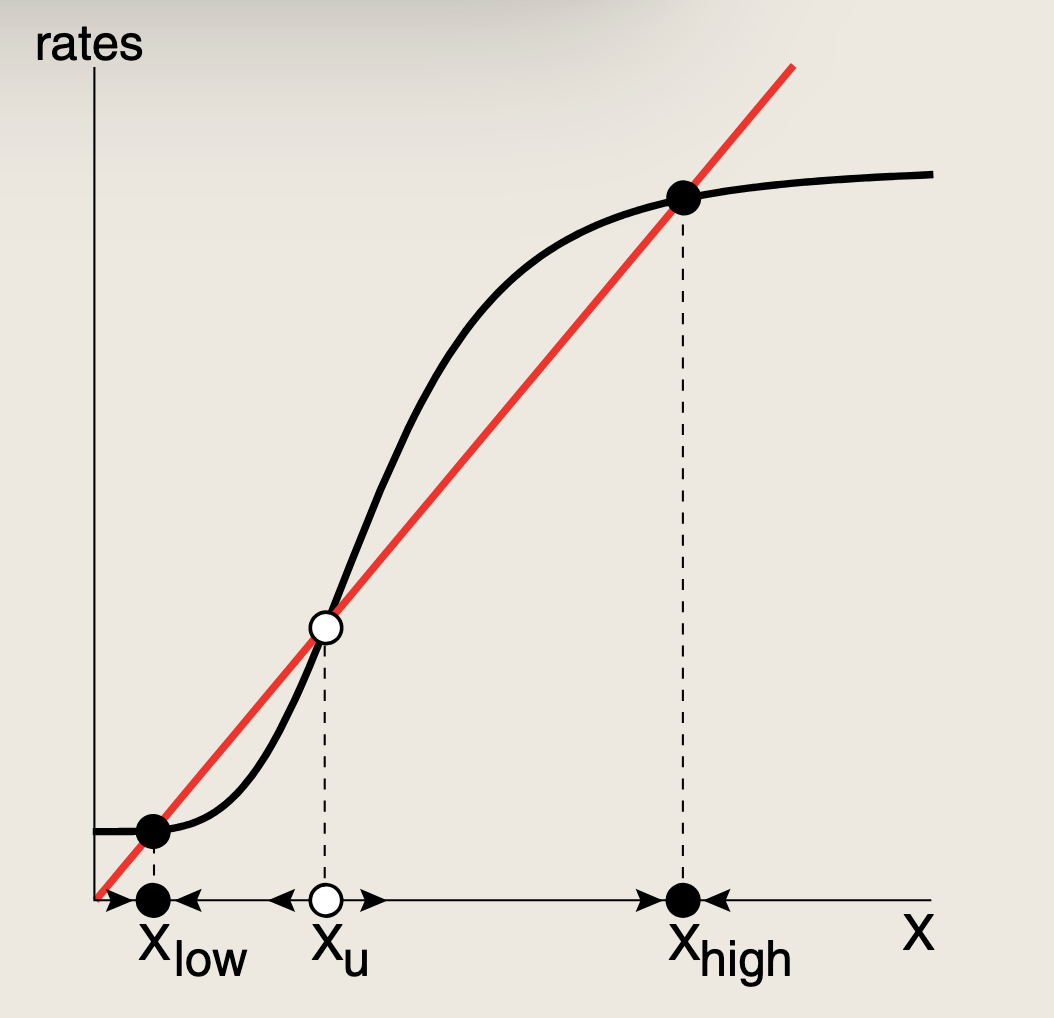
- 三个不动点包括:一个高稳定不动点XhighX_{high}Xhigh,一个低稳定不动点XlowX_{low}Xlow以及一个位于中间的一个不稳定不动点XuX_uXu。
- 中间点是不稳定的,因为将X向其的任一侧稍作移动,X就会远离该点。
- 该系统有两个稳定不动点,这一特性被称为双稳态
- 最终达到哪个稳定状态取决于初始条件:
- 从低值起始,会趋向于XlowX_{low}Xlow
- 从高值起始,会趋向于XhighX_{high}Xhigh
- 我们称这存在两个吸引域,由不稳定不动点分隔
电路记住了它的起始位置。一旦基因被充分激活,它就会锁定在高表达状态,并持续保持开启。
- 假设存在一个转录因子Y也能激活X启动子,X起始于Low,随后Y的激活信号产生,促使更多X生成。X越过阈值XuX_uXu达到高水平,当Y的信号消失后,X仍保持在高水平,即高稳态XhighX_{high}Xhigh。这就是长期记忆
- 长期记忆不是"只要 f(X)f(X)f(X) 是 Sigmoid 就一定出现",而是当正自调控的生产曲线 f(X)f(X)f(X) 足够 S 形、足够陡,并且能和移除曲线 αX\alpha XαX产生三个交点时,才会出现。
此类双稳态电路在发育转录网络中用于做出不可逆的决策,从而将细胞锁定为特定的命运(eg:决定组织类型)
- 当胚胎细胞首次分化为肌肉细胞时,转录因子会激活一个正反馈环,并保持锁定激活状态,从而启动肌肉特异性基因的表达,这些基因负责生成力量所需的肌纤维和蛋白质。正自调控确保细胞不会忘记其肌肉身份
5.1.2 用于决策的双节点正反馈环路
发育网络还呈现出一种网络模体,即两个转录因子相互调控。这种互调控形成双节点反馈环路。
正反馈环路有两种类型:
- 双正反馈环路:两个转录因子相互激活;该环路可以具有两个稳定稳态:
- 在一个稳定稳态下,X和Y均处于开启状态,两个转录因子相互增强彼此生成
- 导致X或Y生成的信号可将系统锁定在开启状态,这种双稳态开关被称为锁定开启机制,因为X和Y要么同时开启,要么同时关闭
- 在另一个稳态下,X和Y均处于关闭状态
- 在一个稳定稳态下,X和Y均处于开启状态,两个转录因子相互增强彼此生成
- 双负反馈环路:两个转录因子相互抑制;该环路同样具有两个稳定的稳态。
- 在一个稳定稳态下,X开启且Y关闭,因为蛋白质X抑制Y
- 另一个稳定稳态相反,X关闭且Y开启
- 双负反馈回路与双正反馈回路不同,它只表达X或Y其中之一,因此该电路也被称为双稳态开关(toggle switch)
- 当由X调控的基因与Y调控的基因属于不同的细胞命运时(eg:分化为不同细胞),该开关机制十分有用
我们在之前可以看到,正自调控也能锁定高表达状态。既然单结点环路足够,为何还要出现双节点反馈环路
- 双节点环路仅在明显延迟后才会锁定,因此可以滤除瞬时的输入信号
可采用零增长线分析法分析上述过程:
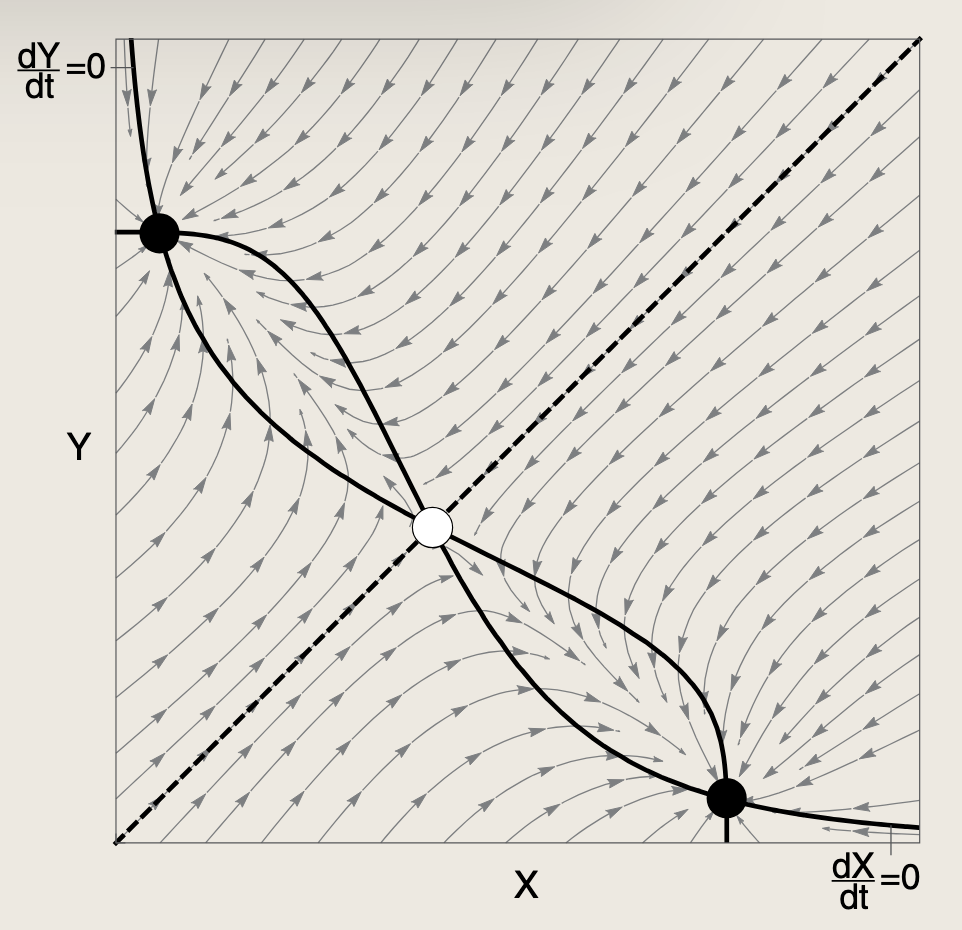
- 双负反馈环路具有双稳态:系统最终会稳定在"X高Y低"或"Y高X低"两种状态之一,中间状态不稳定,因此它适合实现细胞命运的二选一开关。
若两个抑制函数均不具备S形特征,则仅存在单一不动点。此时零增长线的非S形曲线形态组织了它们之间的多次相交。该系统表现为单稳态,X与Y相互抑制,但最终均达到中等浓度且维持不变
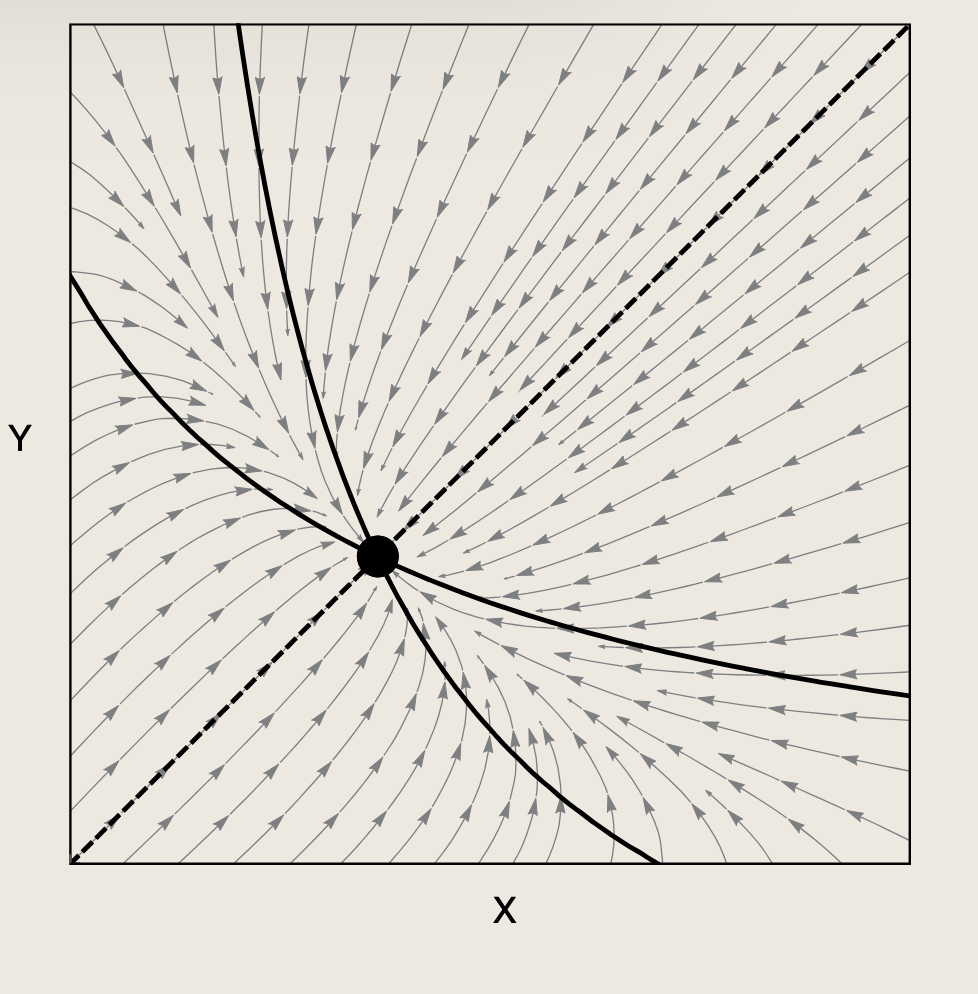
双节点的反馈环路中的转录因子通常还各自具有正自调控作用。正自调控环路可在转录因子积累至一定水平后进一步增强其生成,从而稳定转录因子的"开启"稳态。
5.1.3 调控型反馈与受调型反馈
双节点反馈环路可嵌入发育网络的更大模体中。这类网络呈现出两种主要的三节点反馈环路模体:
- 三角形模式:
- 相互调控的结点X与Y共同调控基因Z,称为调控型反馈
- 最简单的情况下,以双正反馈环路相互激活的X和Y对靶基因Z具有相同的调控符号(均为正或均为负);双负反馈环路往往对Z有相反的调控符号。
- 下图所示的两种符号组合是相干的,即任意两节点之间的两条路径具有相同的总体符号

- 除了调控型反馈模体外,发育网络中还出现一种双节点反馈环路受上游转录因子调控的模体,称为受调型反馈 。同样存在几种相干的符号组合。
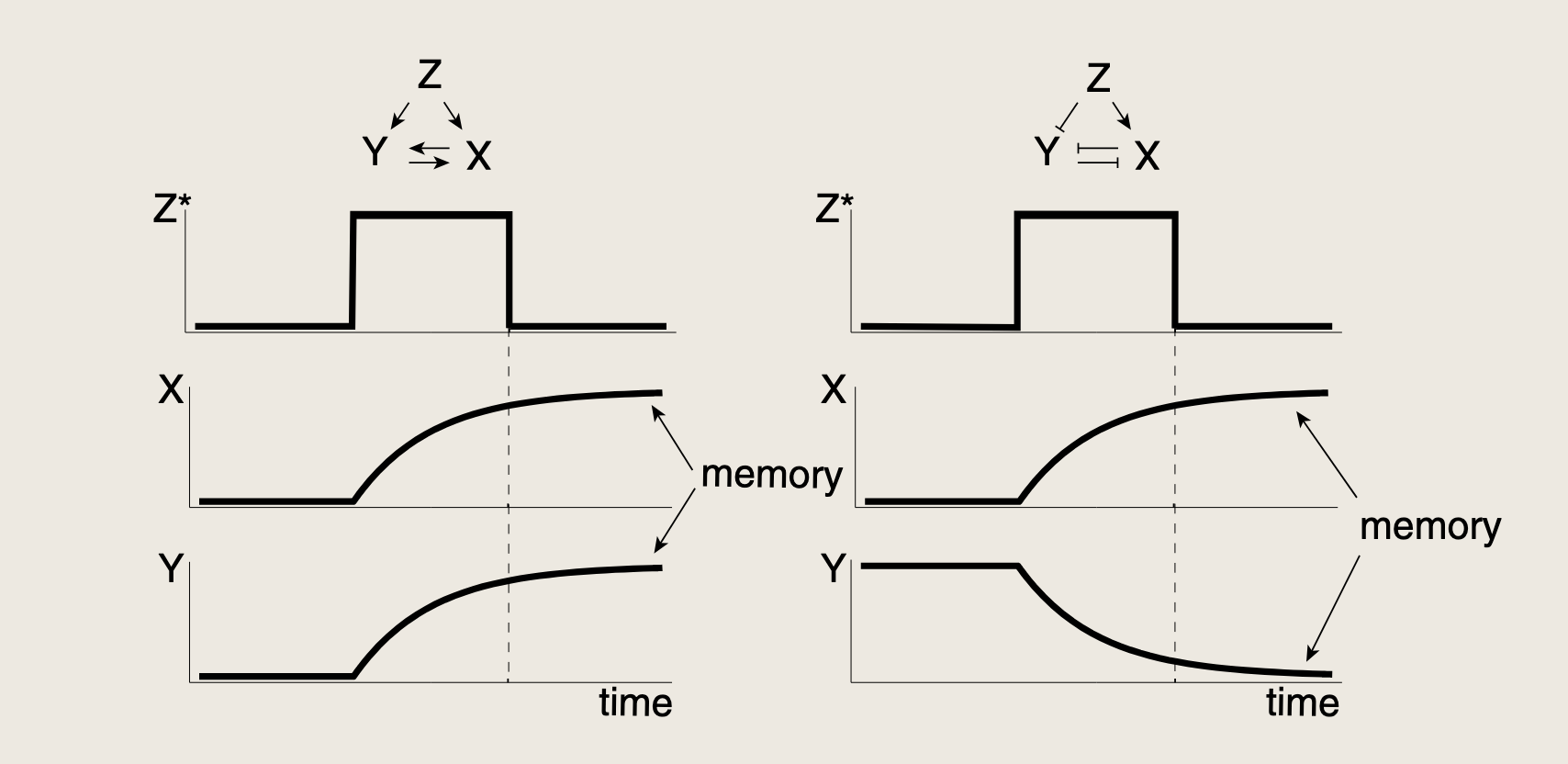
- 对于双促进循环,输入转录因子可以是促进的;对于双抑制循环,输入转录因子可以有不同的作用
- 相互调控的结点X与Y共同调控基因Z,称为调控型反馈
如正自调控PAR一样,受调型反馈模体可被视为一种记忆元件;调控因子Z能够将反馈环路从一个状态切换到另一个状态,使得即使Z失活后改状态仍能持续。因此改电路能够记住Z在过去是否曾处于活性状态。(下层X与Y可以互相维持)这种记忆有助于细胞在决定其命运的最初发育信号消失后,依然维持其原有命运。
5.1.4 长转录级联和发育时序
发育转录网络中另一类重要的模体是长转录级联 。转录级联 是指转录因子X调控Y,Y进而调控Z等一系列相互作用的链条。级联中每一阶段的响应时间由该阶段蛋白质的移除速率决定,即T12=log2(α)T_{\frac{1}{2}} = log_2(\alpha)T21=log2(α)。对于需要快速响应环境胁迫与营养变化的感觉转录网络而言,长级联显得过于缓慢,但是发育网络的工作时间尺度恰好对应于一个或数个细胞世代。
转录级联的时间尺度非常适合引导发育进程。发育过程常采用阻遏蛋白级联,其时序特性相较于激活蛋白级联而言,对蛋白质生成速率的波动具有更强的稳健性。
5.2 蛋白质相互作用中的网络模体
之前提到的感觉型转录网络以及发育型转录网络,它们的运行时间尺度慢至细胞世代时间。为引发快速响应,细胞内还包含更为迅捷的信息处理网络,称为蛋白质相互作用网络(PPI)。PPI之所以快速,是因为它们依靠现有蛋白质之间的相互作用来运作,无需合成新的蛋白质。在PPI网络中,蛋白质通过多种方式相互激活或抑制:彼此结合,进行化学修饰,甚至相互降解,其典型时间尺度为秒至分钟,而转录网络则以小时计。
5.2.1 混合网络模体包含双节点负反馈环路
PPI网络与转录网络以整合方式运作。许多PPI电路以转录因子作为其输出,而PPI网络中的蛋白质则受转录网络调控。整合的网络可描述为具有两类边的网络:
- 一类边代表转录相互作用
- 另一类边代表快得多的蛋白质相互作用
本节将提及一些出现在此类网络中的网络模体,这类网络motif同样可见于整合了超过两个相互作用层次的网络中。
一种非常常见的混合模体是由两个蛋白质构成的反馈环路,它们之间通过两种颜色的箭头相互作用。
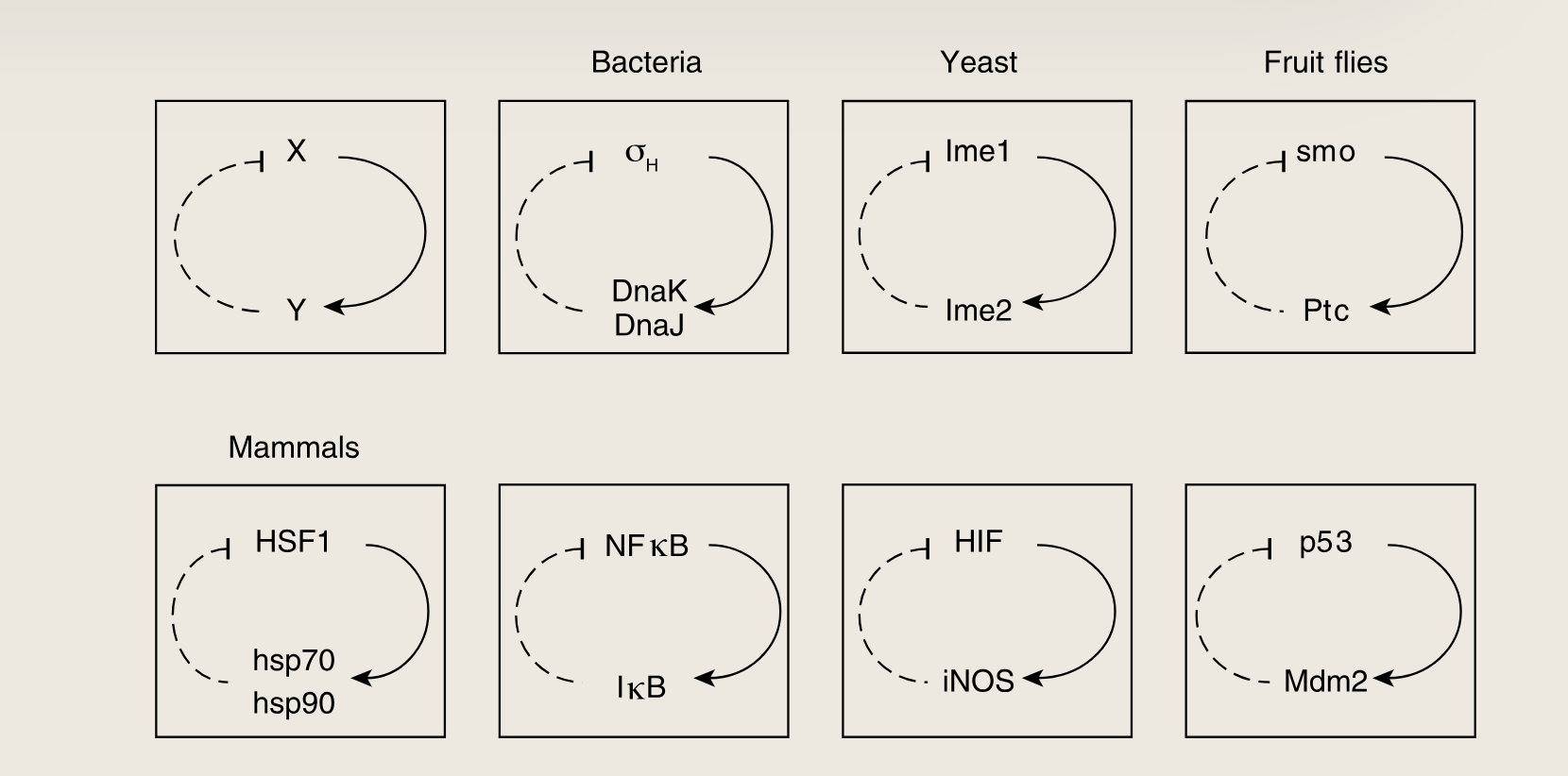
- 在该模体中,蛋白质X是一个转录因子,激活基因Y的转录;而蛋白质产物Y则在蛋白质水平(非转录水平)上与X相互作用,且通常表现为负调控作用 。
- 这种负调控可有多种形式:在某些情况下,Y增强蛋白质X的降解速率;在其他情况下,Y与X结合,通过阻止其解除DNA来抑制其作为转录因子的活性
- 在代谢途径中,最终产物通常负向调控通路酶的转录,以及通路中第一个酶的活性。混合
5.2.2 混合前馈环模体可提供瞬时记忆
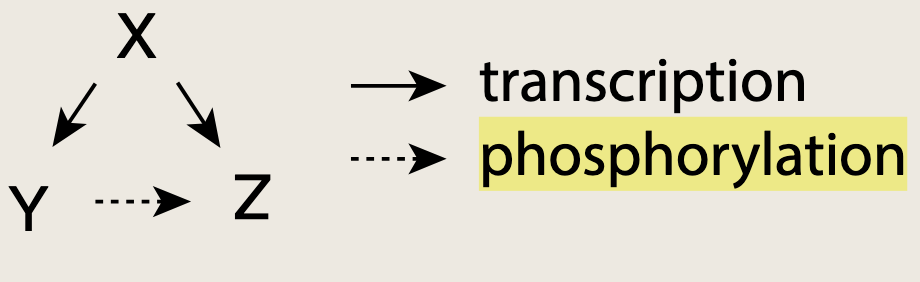
例如X在转录水平激活Y与Z,而Y与Z在蛋白质层面相互作用。这种混合前馈环出现在酵母的交配通路中。在此通路中,输入信号SXS_XSX是另一酵母细胞释放的信息素,输出则是基因Z的表达。
混合前馈环提供了持久性检测功能,信号SXS_XSX必须持续足够长时间,才能使Y蛋白积累并激活Z。该混合前馈环具备瞬时记忆能力;若酵母在近期曾感知过信号SXS_XSX,则它能够对该信号做出更迅速的响应------因为Y蛋白已经存在,无需从头合成便可对Z进行处理。(此处的近期取决于Y的半衰期)
5.2.3 前馈环路展现反馈环路功能的温和版本
| 类型 | 作用过程 | 类似哪种反馈环 | 产生的现象 | 现象解释 | 为什么说是"温和版" |
|---|---|---|---|---|---|
| 一致性前馈环 CFFL | (X→Y),(X→Z),(Y→Z)(X \rightarrow Y),(X \rightarrow Z),(Y \rightarrow Z)(X→Y),(X→Z),(Y→Z) | 类似正反馈 | 延迟响应、短暂记忆 | 输入 (X) 来了以后,(X) 可以直接作用于 (Z),但 (Y) 需要时间积累;如果 (Z) 需要 (X) 和 (Y) 同时存在,短暂输入不会启动 (Z),只有持续输入才能启动。这说明系统能"记住"刚刚是否有持续信号。 | 这种记忆依赖 (Y) 的暂时存在,(Y) 降解后记忆消失,所以只是短暂记忆,不会长期锁定。 |
| 正反馈环 | (X→X),或(X→Y→X)(X \rightarrow X),或 (X \rightarrow Y \rightarrow X)(X→X),或(X→Y→X) | 本身就是正反馈 | 长期记忆、双稳态 | (X) 一旦被外部信号推高,就会继续促进自己或通过 (Y) 促进自己,使系统维持在高表达状态。即使原始输入消失,系统也可能保持 (X) 高表达。 | 这是强版本,容易形成稳定开关,用于细胞命运锁定。 |
| 非一致性前馈环 IFFL | (X→Z),(X→Y),(Y⊣Z)(X \rightarrow Z),(X \rightarrow Y),(Y \dashv Z)(X→Z),(X→Y),(Y⊣Z) | 类似负反馈 | 脉冲、加速响应 | 输入 (X) 来了以后,(X) 先快速激活 (Z),所以 (Z) 上升;随后 (Y) 慢慢积累,并开始抑制 (Z),所以 (Z) 又下降,形成一次"先升后降"的脉冲。 | 它只能产生一次脉冲,因为 (Z) 不会反过来影响 (X) 或 (Y),所以不会来回震荡。 |
| 负反馈环 | (X→Y),(Y⊣X)(X \rightarrow Y),(Y \dashv X)(X→Y),(Y⊣X) | 本身就是负反馈 | 稳定、脉冲、振荡 | (X) 升高后激活 (Y),而 (Y) 又反过来抑制 (X)。如果这个抑制有延迟,系统可能出现"升高---被压低---抑制解除---再升高"的反复过程。 | 这是强版本,如果延迟足够明显,可以产生持续振荡。 |
5.3 神经元中的网络模体
神经元网络与转录网络在网络模体上的相似性令人惊讶,因为两者在空间和时间尺度上截然不同。然而许多模体确是相似的。前馈环同时成为这两种网络的模体。
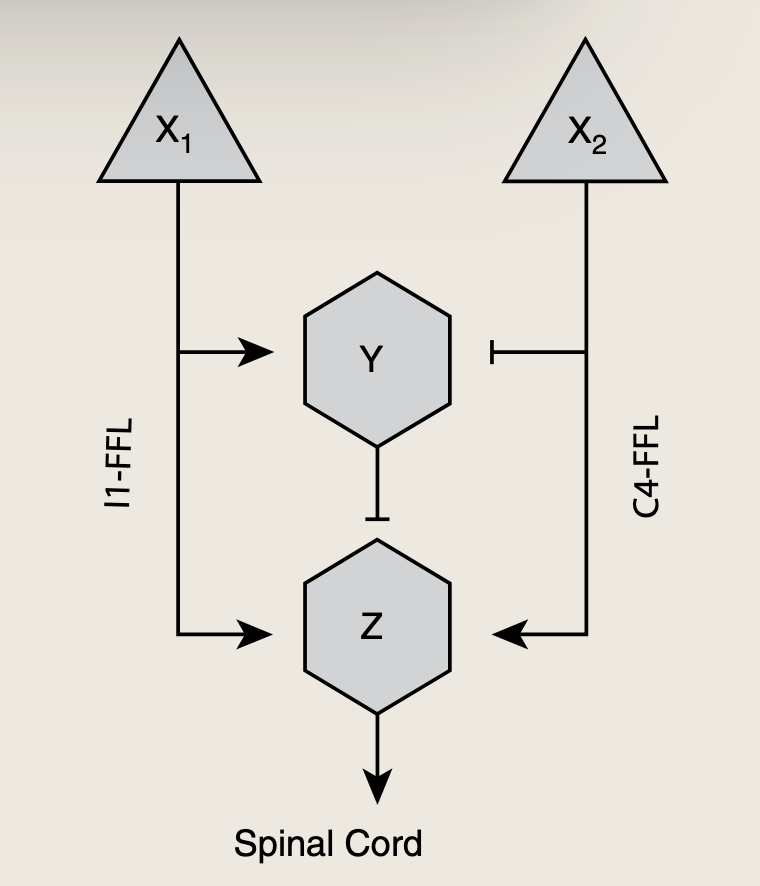
5.3.1 神经元网络中的多输入前馈环
5.3 反思
网络表示掩盖了每个节点和箭头背后的具体机制。然而,正是凭借这种紧凑的描述层次,网络表示有助于凸显网络中不同部分之间以及不同网络之间在回路模式上的相似性。在此分辨率层面,网络动力学可通过简单模型进行分析。我们只关心X激活或抑制Y,而不精确关注其在生化层面如何实现。这种抽象使我们能够将网络模体定义为每种网络的功能性构建模块。这些构建模块通常以允许根据每个单独模体的动力学来理解整个网络动力学的方式连接在一起。因此,无论是局部模式层面,还是将这些模式组合成更大回路的层面,生物网络似乎都展现出一定程度的简洁性。
Chapter 6 如何构建生物振荡器
6.1 震荡需要负反馈和延迟
振荡器的核心是一个负反馈回路 。分子X的作用是降低自身浓度,从而使其从高水平降低至低水平,再回升至高水平,循环往复。然而仅凭负反馈本身并不足以产生振荡。负自反馈并不能造成振荡,而是不断的维持稳态(负自反馈几乎是"及时纠错"的,系统不会明显冲过头)
生物电路中的延迟可通过在负反馈回路中增加组件,从而延长回路路径长度来实现。
自调控仅仅是一条单步路径,即一个闭合箭头,再增加一个物种Y,便得到由两个箭头组成的负反馈回路。
可以隐约看到阻尼振荡:
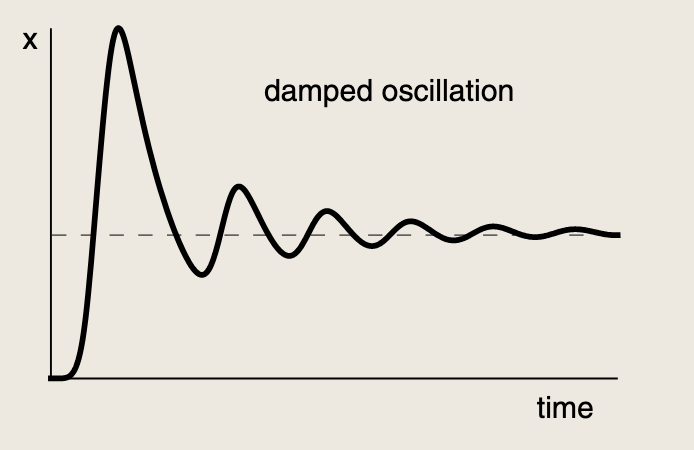
阻尼振荡需要满足两个条件:
- 强反馈,最好具有协同性
- 两个箭头的时间尺度相近。如果相差悬殊,其中一个箭头远快于另一个,则快速路径无法成为有效的延迟元件,电路呈现过阻尼状态,效果实际上等同于无阻尼振荡的自调控。
已解示例6.1:证明如果时间尺度足够相似且反馈足够强,则两个分量负反馈环路表现出阻尼振荡


此类线性方程的解是指数时间函数的和:C1eλ1t+C2eλ2tC_1e^{\lambda_1t}+C_2e^{\lambda_2t}C1eλ1t+C2eλ2t,指数中的λ1\lambda_1λ1和λ2\lambda_2λ2是矩阵的两个特征值
- 当特征值具有虚部时,便会出现阻尼振荡
- 根据欧拉公式ea+ib=ea(cos(b)+isin(b))e^{a+ib} = e^a \left(\cos(b) + i\sin(b)\right)ea+ib=ea(cos(b)+isin(b)),解表现为一个震荡波,其频率由虚部(ω=2π/b\omega = 2\pi / bω=2π/b)决定,而振幅在实部为负(a < 0)时以指数形式衰减至0.该结果成为螺旋不动点,系统会呈螺旋状收敛值稳态解。
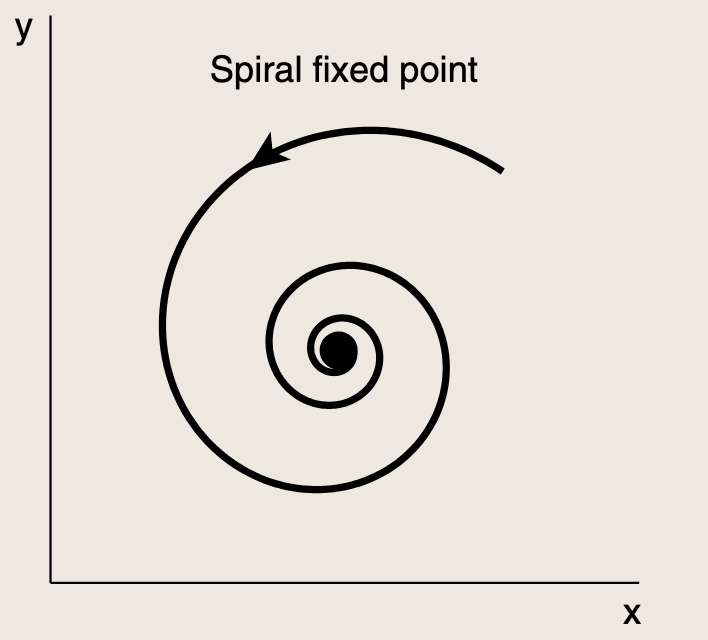
为了解特征值,我们需解矩阵J的特征方程:
- (−α1−λ)(−α2−λ)+β1β2=0(-\alpha_1 - \lambda)(-\alpha_2 - \lambda) + \beta_1\beta_2 = 0(−α1−λ)(−α2−λ)+β1β2=0
得到特征值方程:
$2\lambda_{1,2}
-(\alpha_1+\alpha_2)
\pm
\sqrt{(\alpha_1-\alpha_2)^2 - 4\beta_1\beta_2}$
由于负移除项的存在,该电路的特征值实部始终为负,因此稳态稳定,所有初始条件都会留回稳态。
之前提到过,阻尼振荡发生在特征值具有虚部时,即平方根内的项均为负值:
- (α1−α2)2<4β1β2(α_1−α_2)2<4β_1β_2(α1−α2)2<4β1β2
- 右侧项称为反馈强度,当两条分支的时间尺度相等(α1=α2\alpha_1=\alpha_2α1=α2)时,无论反馈强度大小如何,阻尼振荡均会出现
时间尺度的差异 ∣α₁ − α₂∣ 越大,产生阻尼振荡所需的反馈强度 β₁β₂ 也越大。若反馈强度不足以弥补时间尺度的分离程度,系统将呈过阻尼状态,单调衰减至不动点,无任何超调现象。因此,时间尺度的高度分离会抑制振荡趋势。
反馈强度 β₁β₂ 由调控函数 g 和 f 在稳态点处的斜率 dgdX\frac{dg}{dX}dXdg和 dfdY\frac{df}{dY}dYdf决定。这些调控函数越陡峭(例如希尔系数越高),反馈强度就越大。因此,协同性会增强振荡的倾向。
6.2 噪声可在理论上仅存在阻尼振荡的系统中诱发持续的振荡
当噪声足够强时,它便会使系统偏离螺旋不动地啊,从而阻止振荡的衰减。
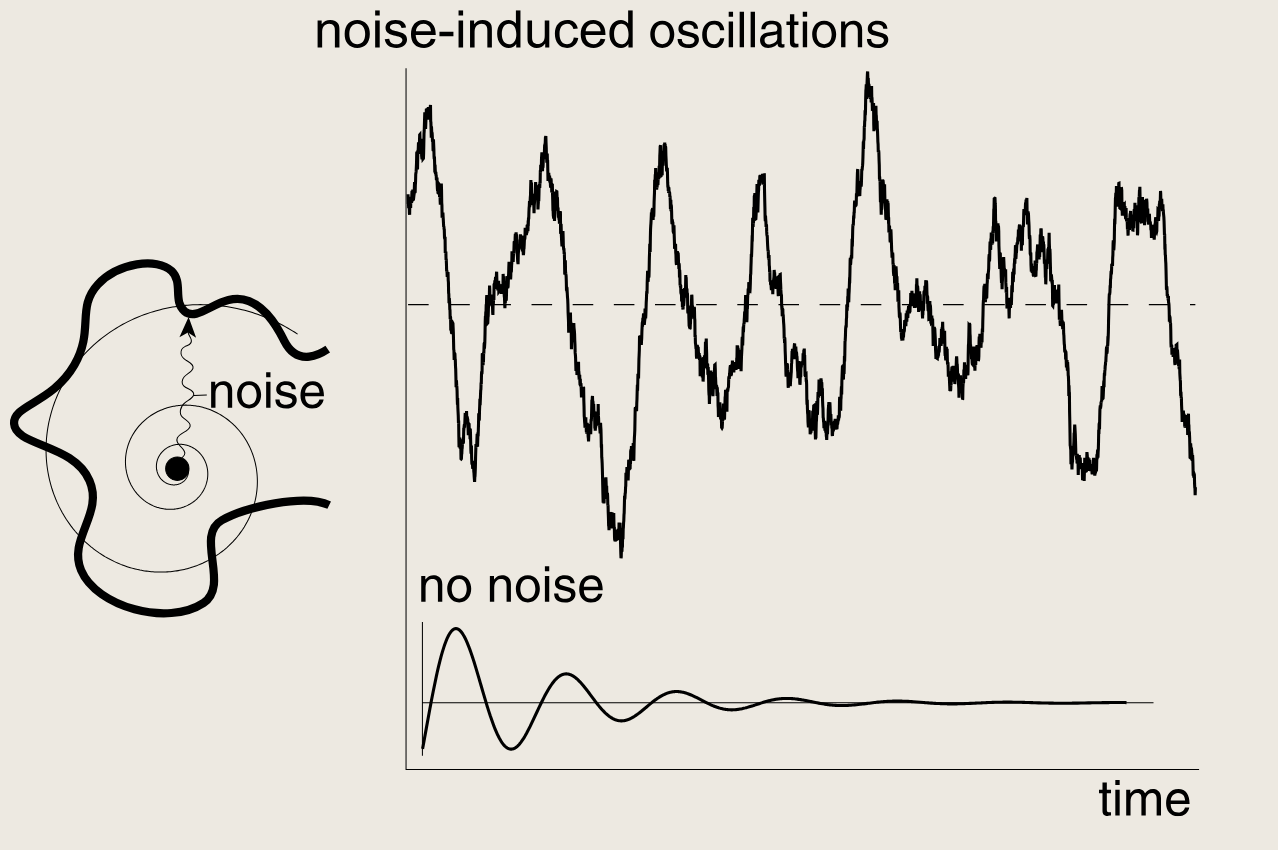
6.3 延迟振荡器
当反馈回路由三个或更多步骤构成时,即使在没有噪声的情况下,也能产生完全成型的无阻尼振荡 。此类回路要实现振荡,必须具备强反馈 ,且各步骤的时间尺度相近。
将三种抑制子的基因及其调控区域组合在大肠杆菌内,使美中抑制子依次抑制循环中的下一个抑制子。
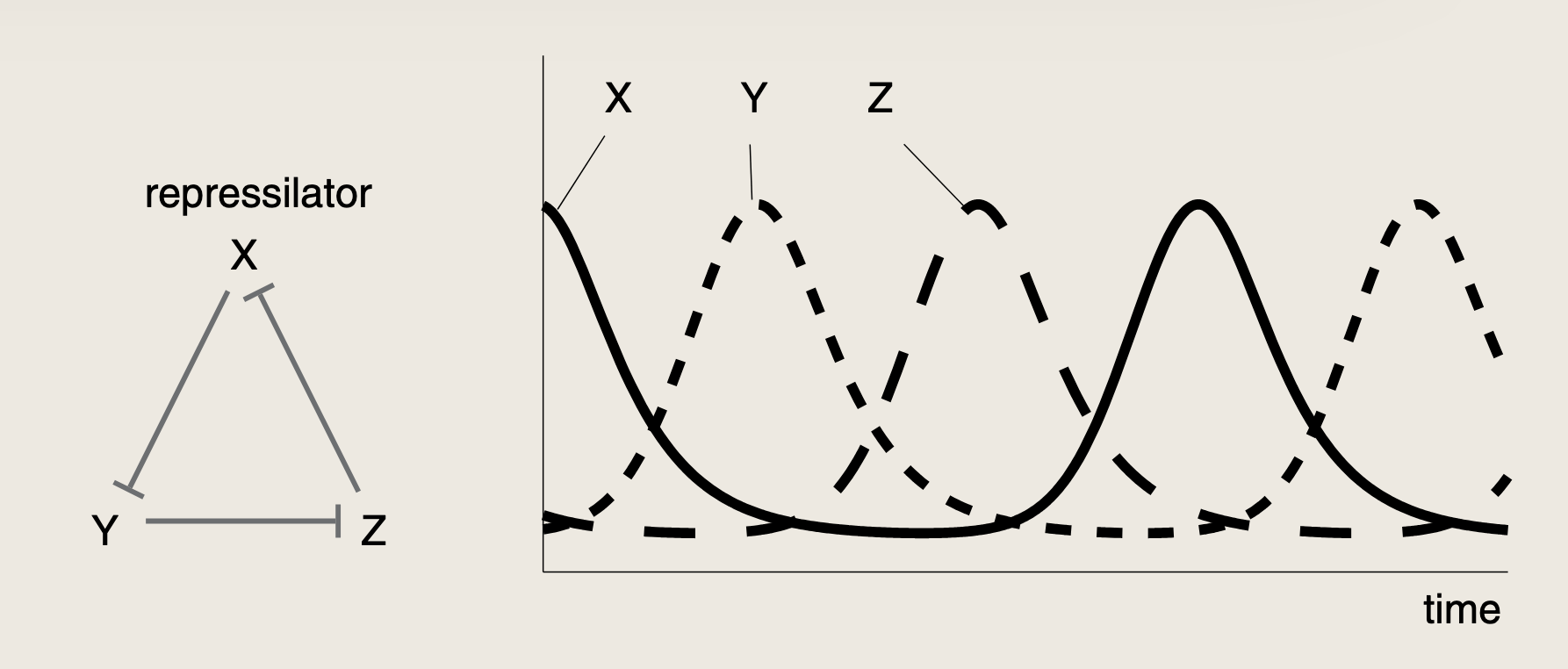
回路中的组件越多,振荡所对应的参数范围就越广泛,而实现振荡所需的协同性程度越低。
6.4 需要生物振荡器有耦合的正负反馈环路模体
综上所述,负反馈结合延迟 和/或 噪声可以产生振荡。非线性(协同性)以及相对臂之间相似的时间尺度有助于反馈回路产生振荡。
当我们审视生物学中研究最充分的振荡器电路时,会额外发现一个特征------在负反馈环路的基础上增加了一个正反馈环路。
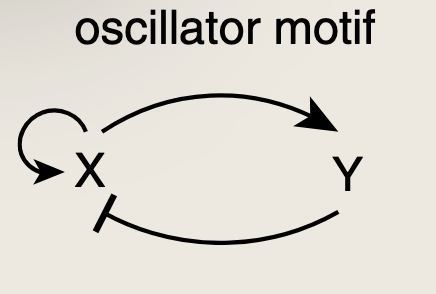
正反馈增加了延迟,延迟有助于振荡,扩展了能够产生振荡的参数范围。(正反馈可以使双节点负反馈环路即使在一个臂远快于另一个臂的情况下也能表现出持续振荡)。
没有正反馈时,两个特征值具有负实部,我们得到不动点或稳定螺旋。正反馈可以使实部变为正值,将稳定螺旋转变为不稳定螺旋。轨迹呈螺旋状向外扩散。
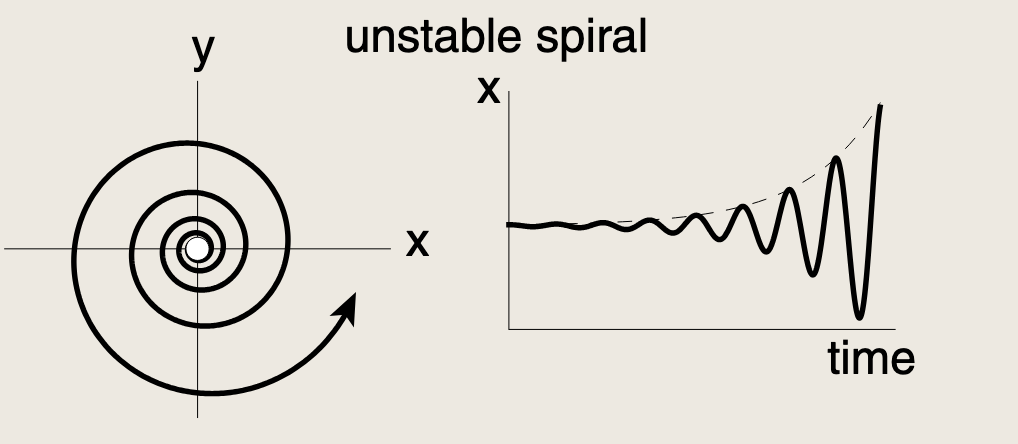
轨迹不可能无限发散。一旦浓度上升到足够高,所有反馈项都会饱和,我们只剩下移除项来将浓度推回。这种受限的轨迹会最终稳定在一种持续振荡中,称为极限环:
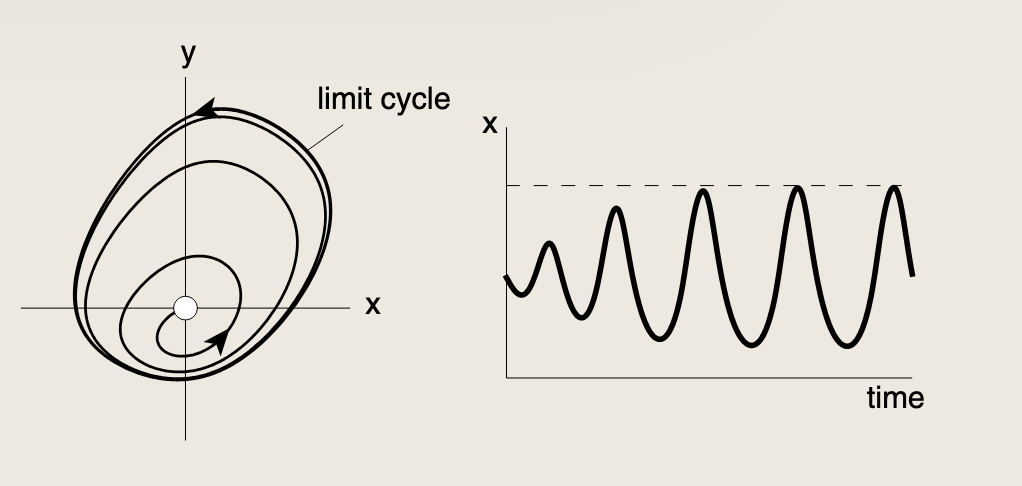
6.5 利用双正反馈环路实现鲁棒双稳态
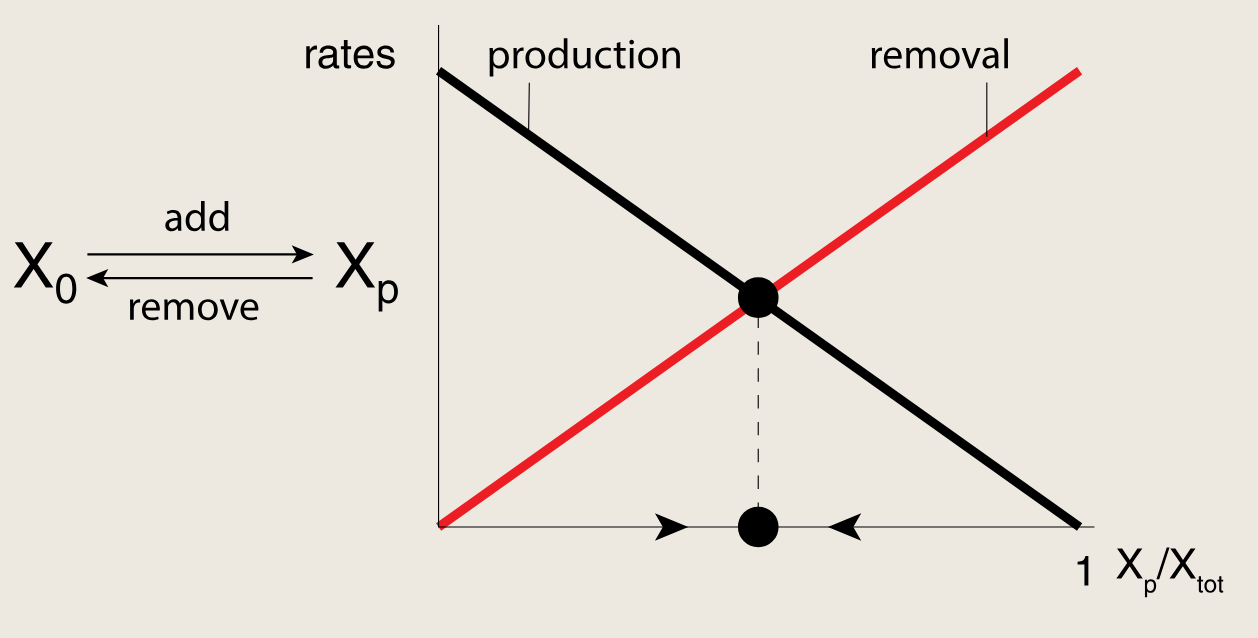
XpX_pXp通过两种方式提升自身水平:
- 激活自身生成
- 抑制自身向X0X_0X0的转变
这形成了两个正反馈环路
为何需要两个环路而非一个?双环路电路能使双稳态在参数大幅度变化的同时保持鲁棒性
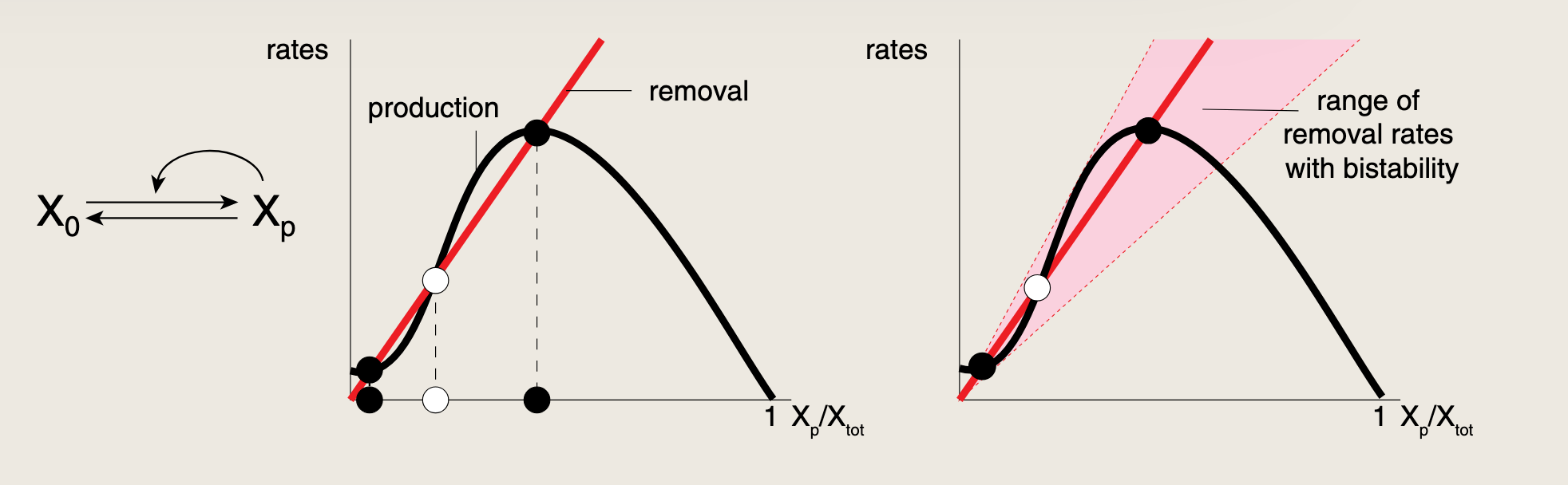
如上图(右)所示,这是一个无反馈的电路,仅包含有激酶催化的X0X_0X0磷酸化生成XpX_pXp,以及由磷酸酶催化的XpX_pXp去磷酸化回到X0X_0X0的过程。
- 移除速率为一条上升直线,即XpX_pXp乘以磷酸酶的速率
- 生成速率为一条下降直线,即X0X_0X0乘以激素酶的速率,当所有X均以XpX_pXp形式存在时降至0,因为再无X0X_0X0可供磷酸化
- 两条直线相交的点即位生成等于移除的稳态点;此时仅有一个焦点,对应单一的稳态,无双稳态。
现在加入一个正反馈环路,使XpX_pXp激活自身的生成,该环路可展现双稳态,但这种双稳态非常脆弱,一旦移除速率参数发生微小变化便会消失。
- production曲线之所以是驼峰形:
- 一开始XpX_pXp很少,正反馈还没启动,生成速率低
- 随着XpX_pXp增加,XpX_pXp促进自己的产生
- 当XpX_pXp接近XoutX_outXout时,几乎所有X都已经是XpX_pXp,剩下的X0X_0X0很少了,没有足够的底物可以转化,所以生成速率又下降,最后为0
相比之下,双正反馈环路意味着XpX_pXp既可以增强自身生成,又可以减少自身消除。这使得生成曲线和消除曲线呈现对称性,两者均为驼峰型,因此其交点对参数变化的敏感度大幅降低。提供了鲁棒的双稳态,进而产生稳健的松弛振荡。
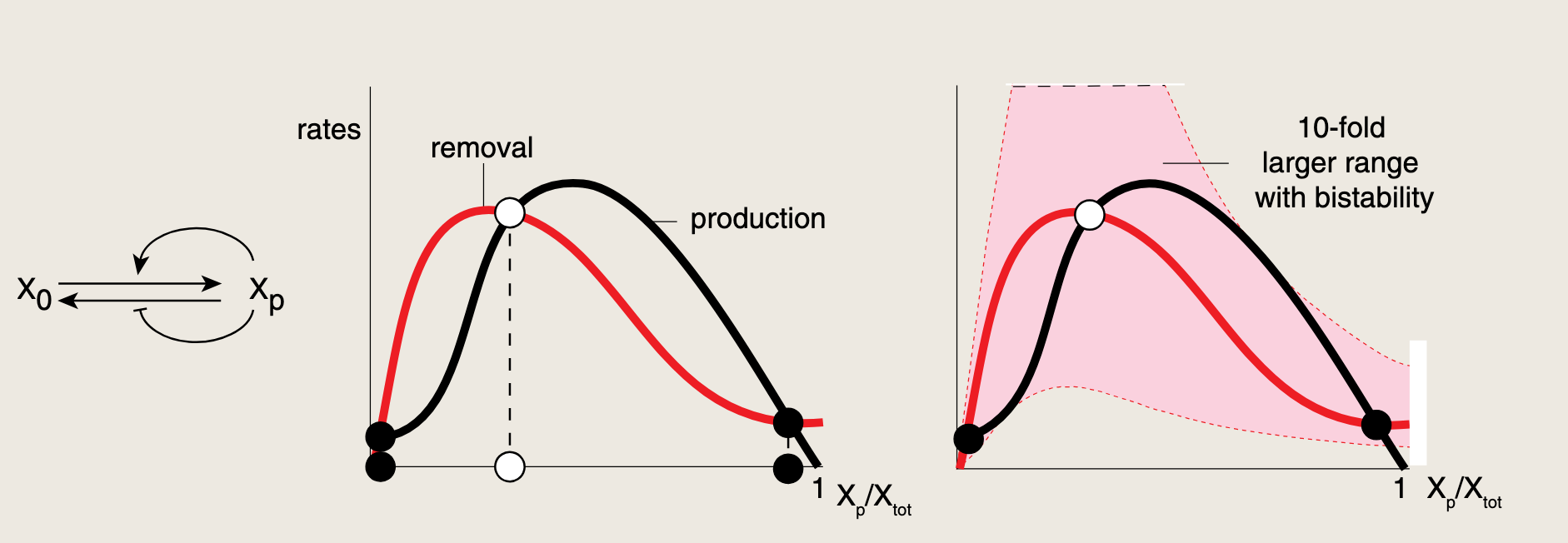
- XpX_pXp越多,越容易继续变多,也越不容易变少
总之,振荡需要负反馈,延迟与协同性可促进振荡。噪声可以将阻尼振荡转为持续脉冲
许多生物振荡器都采用一种模体,即负反馈环路与正反馈环路耦合,产生频率可调,振幅文件的尖峰状脉冲