论文总结
一、研究背景与现存缺陷
- 多组学(mRNA、CNV 等)是精准医疗核心数据,传统深度学习无法充分利用基因互作、蛋白互作等图结构先验生物知识。
- 现有 GNN 多组学方法仅使用单一生物先验图,无法融合多源先验信息,小样本 / 少基因特征场景性能差。
- 多视图 / 多组学模型缺少对齐多先验图谱表征的机制,信息互补性挖掘不足。
二、核心创新(MPK-GNN)
- 首个融合多生物先验图 + 对比学习的 GNN 多组学框架,端到端可拓展。
- 四大模块化结构:
- 特征层模块:共享 GCN,基于多先验图学习基因层面表征;
- 投影模块:对比学习对齐多张先验图表征,最大化一致性;
- 样本层模块:MLP 提取患者全局组学特征;
- 任务模块:拼接多级表征适配下游分类等任务。
- 联合损失:分类交叉熵损失 + 监督对比损失,λ 平衡两项权重。
- 输入三类生物先验图:GGI 基因互作、PPI 蛋白互作、CoExp 共表达网络。
三、实验设置
- 数据集:TCGA 泛癌多组学、BRCA 乳腺癌亚型数据集;按基因数量划分 TCGA-m 子集。
- 对比基线:传统机器学习 (SVM/RF/KNN)、多视图模型、主流多组学 GNN (MOGONET/CMSC)。
- 验证任务:癌症分子亚型分类、乳腺癌亚型识别。
四、关键实验结论
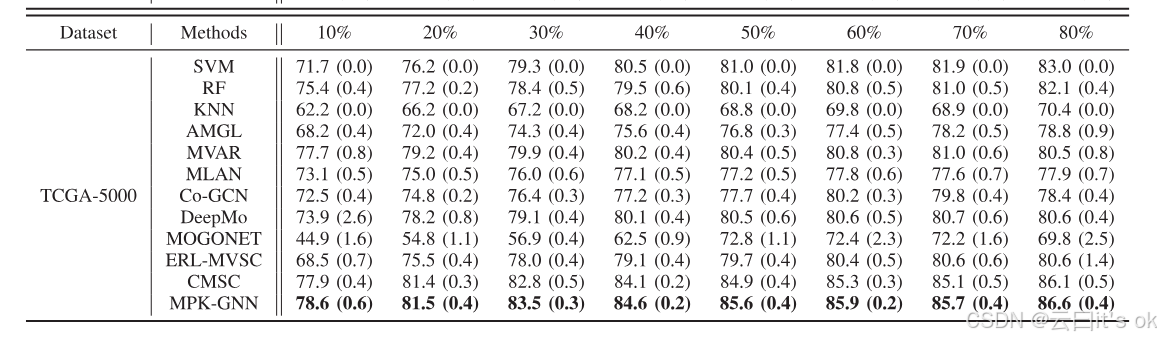
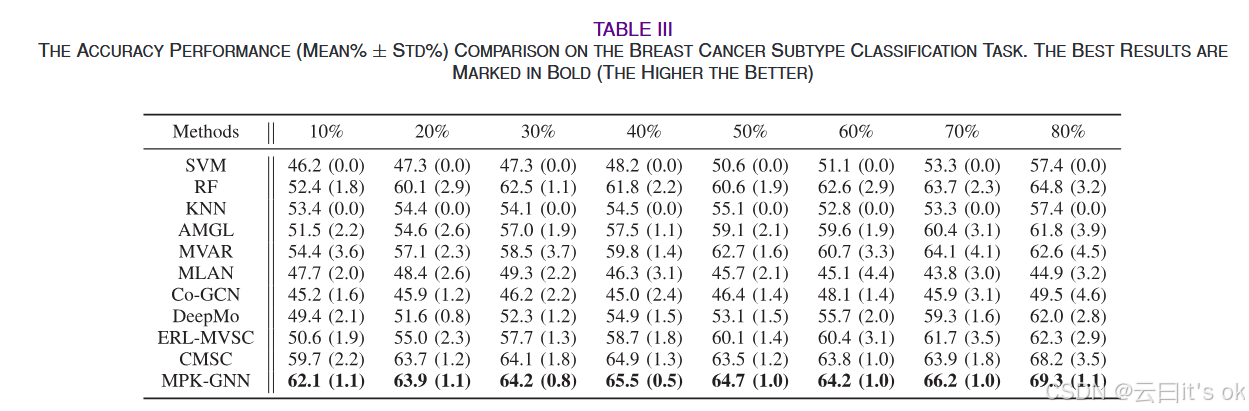
- 性能优势:MPK-GNN 在全部数据集、不同训练样本占比下准确率最优,方差更小、鲁棒性更强;小样本 / 少基因场景提升最显著。
- 多先验有效性:融合全部 3 种先验图效果远优于单种先验 / 无先验基线;CoExp、GGI 优于 PPI 网络。
- 对比学习增益:移除投影对比分支后模型精度显著下降,共享结构同时减少参数量。
- 混淆矩阵:混合分子亚型(多癌起源)最难区分,为模型主要错分样本。
- 超参鲁棒:对比损失权重 λ、温度系数 β 大范围调整下性能稳定;Bonferroni-Dunn 检验证明性能显著优于绝大多数对比方法。
五、结论与未来工作
- 总结:多先验 + 对比学习 GNN 能高效挖掘多组学生物先验,适配小样本精准医疗任务,模块可灵活拓展至各类多组学任务。
- 未来优化:改进样本层模块、在更多多组学任务上验证模型。
摘要
随着生物技术的发展,精准医疗收集了大量多组学数据。存在多项基于图形的生物学知识,涉及组学数据,例如基因-基因相互作用网络。近年来,将图神经网络(GNN)引入多组学学习的兴趣日益增加。然而,现有方法尚未完全利用这些图形先验,因为没有方法能够同时整合来自多个来源的知识。为解决该问题,我们提出了一个多元组学数据分析框架,将多项先验知识整合进图神经网络(MPK-GNN)。据我们所知,这是首次尝试将多张先前的图表引入多组学数据分析。具体来说,该方法包含四个部分:(1)一个特征级学习模块,用于从之前的图表中聚合信息;(2)通过优化对比损耗,最大化先行网络间的一致性的投影模块;(3)样本级模块,用于从输入多组学特征中学习全局表示;(4)针对任务的模块,可灵活扩展MPK-GNN以适应各种下游多组学分析任务。最后,我们验证了所提出的多组学学习算法在癌症分子亚型分类任务中的有效性。实验结果显示,MPK-GNN优于包括多视图学习方法和多组学集成方法在内的其他先进算法。
引言
精准医学是未来医疗的重要组成部分,因为它为患者提供个性化治疗,从而改善治疗效果并降低成本。例如,由于复杂的临床、病理和分子特性,同一治疗对乳腺癌患者可能有不同的效果1。随着高通量生物医学技术的发展,疾病特征可以通过多种基因组学来表述。例如,癌症基因组图谱(TCGA)数据集包含mRNA表达、DNA甲基化和拷贝数变异(CNV)数据。与单组学方法相比,多组学方法能够在多个数据中捕捉一致且互补的信息,构建准确且富有洞察力的模型2。因此,有必要引入多组学数据,应用于精准医疗的不同任务,涵盖药物反应预测3、基因发现4到生存分析5。深度神经网络(DNN)因其强大的表示能力,已被成功应用于自然语言处理6和计算机视觉7等多个领域。近年来,越来越多的研究人员结合多组学数据和DNN的优势来解决各种精准医疗问题8、9、10。具体来说,基于DNNs的方法通常包含三个阶段:(a)将高维遗传特征转化为高层次语义嵌入;(b)从多重嵌入中学习统一表示;(c) 将学习到的表示应用于下游任务。然而,已有若干基于图结构的生物学知识,如蛋白质-蛋白质相互作用网络。传统的深度学习范式很难有效利用这些数据。近年来,已引入多种图神经网络(GNN)用于处理非欧几里得数据,如图和流形,并在多个领域取得了成功11、12、13。例如,Wang 等人14提出了一种基于图卷积网络的多组学方法,包含针对组学的学习和跨组学相关学习,用于生物医学分类。此外,Li等人15设计了一个基于GNNs的计算模型,结合一个基因相互作用网络,用于癌症分子亚型分类。然而,这些方法要么不使用先验的生物图数据,要么只使用其中一种。因此,之前的生物图未能被充分利用来提升计算模型的性能。
为解决上述问题,我们提出了一种新的端到端学习框架,将多项先验知识整合进图神经网络(MPK-GNN)进行多组学数据分析。该框架由四个模块组成:基于特征的学习模块、投影模块、基于样本的学习模块以及任务特定模块,如图1所示。基于特征的模块是一个共享图卷积网络,利用多元组学特征和多项先验知识作为输入,学习特征级表示。然后,将学习到的局部嵌入作为共享投影函数的输入,通过优化对比目标来最大化它们之间的一致性。注意,基于特征的模块和投影模块构成了一个共同的对比学习框架。此外,基于样本的模块设计用于为每个输入样本学习全局嵌入。最后,特征级嵌入和样本级表示被串接并作为输入到任务特定模块中。整个框架采用包含对比损失和任务特定优化损失的联合目标进行优化。具体来说,本研究解决了癌症分子亚型分类任务及其在乳腺癌中的应用。本工作的主要贡献总结如下:我们提出了一种新的可扩展端到端深度学习框架,用于学习各种多组学数据分析任务的多层次表示。据我们所知,这是首次采用对比学习框架,将多个先验知识同时纳入GNN。实验结果显示,MPK-GNN在一个流行的TCGA数据集上相比最先进的方法取得了显著提升。本文结构如下。第二部分简要回顾了相关文献,包括GNN、多视角学习和多组学分析。随后,第三部分展示了所提议的MPK-GNN框架的详细内容。我们在第四部分评估和分析MPK-GNN的疗效。最后,结论和可能的方向见第五部分。
相关工作
本节回顾了多组学数据分析和图神经网络的几种相关方法。我们还简要比较了MPK-GNN与现有方法。最后,我们介绍了几种流行的多视图学习方法。
多组学分析
与单组学方法不同,多组学数据分析算法旨在有效整合多个输入特征以实现下游任务,并且已被广泛探索4、16。近年来,引入了多种深度学习范式用于多组学数据分析,以学习低维表示5、17。Chaudhary等人8提出了一个多层自编码器框架,用于从三种组学数据中学习紧凑嵌入,用于肝细胞癌存活预测。Sharifi-Noghabi 等人3 提出了基于 DNNs 的多组学集成模型,该模型由多个特定组学编码子模块和一个联合表示学习模块组成。为了区分泛癌转移的现状,Albaradei 等人9基于三种基因组学数据设计了一种具有三层异质性的卷积变分自编码器。此外,Zhang等人10开发了一个统一的多任务深度模型,包括表示学习模块和任务专用模块,用于多组学数据分析任务。尽管上述多组学学习方法实现了相当的性能,但它们通常仅使用多组学特征,无法利用生物先验提升成果的知识。相比之下,提出的基于GNNs的框架适合进一步利用多项先验知识,以提升计算模型的鲁棒性和性能。
图神经网络
近年来,由于存在多种图结构化数据,如蛋白质-蛋白质相互作用网络,已有多项研究为生物信息学领域引入了图神经网络 18, 19。此外,一些研究者致力于将各种图网络应用于多组学数据分析14、15。Liu等人20设计了一个癌症药物反应计算模型,由多个子模块和一个统一的图卷积网络组成,基于癌细胞的多组学谱。Peng 等人21在异构图上应用卷积图,以了解细胞系和药物的潜在表示,随后构建了癌症细胞系-药物相关矩阵用于预测药物反应。同样,Feng等人13对构建的多边图进行了消息传递以预测药物反应。对于分子亚型分类,Yin等人22提出了基于GNNs的框架,通过聚合相关子图的信息,学习每个样本的新表示方式。MPK-GNN与上述方法的区别在于,该方法结合了对比学习目标与图卷积网络,以同时利用多个先验图。因此,这些先前的图让MPK-GNN更加稳健,尤其是在只有少数监督样本和组学特征可用时。
多视图学习
多视图学习是机器学习中的一个热门话题,已有多种方法被提出用于各种视觉和语音任务23、24。多视图学习方法的目标还包括捕捉多个输入之间的一致性和互补性信息。因此,将其应用于多组学分析任务具有前景。例如,Nguyen 等人25展示了多视图方法在学习样本异质性和揭示串扰模式方面的优势。本研究还比较了包括传统机器学习和深度学习范式在内的多种多视图学习算法与所提出的多组学集成框架。
所提出的方法
本节介绍了提出的多组学学习框架。MPK-GNN 包含四个子模块:特征级嵌入学习模块、投影模块、样本级表示学习模块和任务特定模块。关于这些子模及优化目标的进一步细节将在下文详细说明。
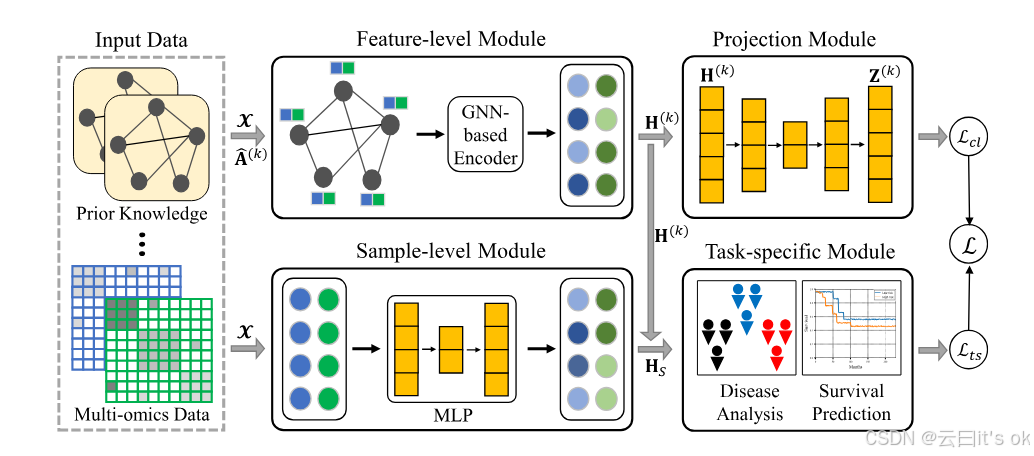
图1。拟议多组学学习框架概述。基于特征的模块汇总输入特征的信息,根据前述图表生成特征级嵌入。投影模块用于重建每个先验知识的表示。相比之下,基于采样的模块由多层感知器(MLP)实现,用于学习采样级表示。然后,这些多层次表示被串接用于下游任务。最后,所提出的MPK-GNN框架通过组合目标函数L进行了优化。
预定义
多组学数据分析旨在学习从多个输入特征 X = {X(v)}vV=1 中对下游任务进行全面的嵌入。这里,X(v) ∈ Rn×m, v = 1, 2, ... . . , V 代表第 v 个组学特征。n、m 和 V 分别表示样本数量、基因和组学。基于输入基因,根据现有知识数据库(如BioGrid 26和STRING 27)生成多个先前图G(k) = (V, E(k)), k = 1, 2, ... . , K这里,节点集 V 代表所选基因,E(k) 代表第 k 个先验知识的边集,K 代表先验知识图的总数。基因之间的关系是按照某些规则建立的。例如,具有强归一化相关性(>0.6)的蛋白质间相互作用将被加入边集。注意,构建的 G(k) 不同于其他域中使用的,因为节点集 V 的元素是遗传特征而非样本。因此,信息聚合机制被应用于特征层面。基于构造的先验知识图G,可以根据规则Aij = eij构建对应的邻接矩阵A∈Rm×m。具体来说,如果存在边,eij = 1,则存在 vi 和 vj 之间,否则 eij = 0。为了减少深度网络中梯度消失或问题爆炸的影响,本研究中使用了A的重整化技巧形式。具体来说,精炼形式表述为:
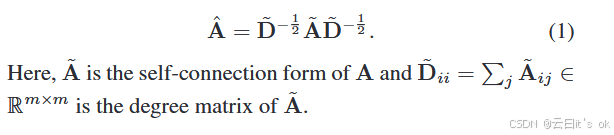
MPK-GNN的方法论
在生物信息学处理中,存在若干基于图的先验数据,如基因间的相互作用。这些先前的图表可以提供有用的知识,帮助提升学习性能并保持学习算法的鲁棒性,尤其是在信息有限的情况下。因此,将这些先有知识同时引入多组学学习方法非常重要。共享架构:借鉴28中的数据增强,我们设计了一个基于GNN的共享对比学习架构,该架构由投影模块和基于特征模块组成,通过最大化先前图之间的一致性来学习紧密对齐的表示。特征级模块包含多层图卷积编码器、最大池化层和平面化层。这里,我们表示共享图编码器为fG(XG, Aˆ (k); Θ),其中XG ∈ Rm×nV 表示通过重塑输入多组学特征X来表示特征级模块的输入。此外,ˆA(k) 是第 k 个先验图的传播矩阵,由 (1) 得到,Θ = {W(l) G , b(l) G }lL=1 表示图编码器的可训练共享权重和偏置,L 表示层的总数。因此,fG 在第k个前图上的第l层计算如下:

注意,每个先验知识的最后一层输出将被重新塑造为 H(k) G ∈ Rn×dL×m 作为下一个子模,其中 dL 表示训练表示的维数。消息传递阶段后,为学习到的嵌入 H(k) G 应用图的最大池层,以降低维度并形成更紧凑的表示。这里,最大池化层表示如下:

投影模块旨在为每种先验知识重建一个表示。我们通过浅神经网络实现该模块,记为fP (H(k); Ω),其中Ω = {W(t) P, b(t) P }tT=1和T分别是可学习的共享参数和层数。第k个先行图的t层输出计算为:

基于样本的模块:我们引入基于样本的并行模块,以学习每个样本的全局表示。该模块由多层感知器 fS(XS; Ψ)实现,其计算过程如下所示:
任务特定模块:基于上述模块,我们设计了一个任务特定模块,可根据目标任务由不同网络实现。学习到的全局特征HS和所有局部特征{H(k)}kK=1被串接起来,形成该模块的统一输入:
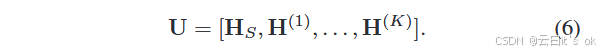
这里,每个样本表示为 (dS + dF K) 维向量。以癌症分子亚型分类为例,我们将U输入全连通网络,随后使用softmax函数,以获得标签的概率分布矩阵。
MPK-GNN的训练
MPK-GNN通过结合对比学习损失Lcl和任务特定学习目标Lts进行优化。对于Lcl,我们将所有重建的表示{Z(k)}kK=1叠加成一组节点表示Z,行数为nK,并用f∈F≡{1, . . . , nK}表示每个节点的索引。然后,对比损耗Lcl的形成方式如下:
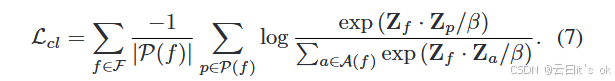
这里,Zf 表示 Z 的第 f 行,β 是尺度超参数,P(f ) 是指示样本与锚点 vf 具有相同标签的索引集合。此外,A(f) ≡ F\{f } 表示除目标节点 vf 外所有样本的索引集合,且 ·表示内积(点积)。对于无监督学习任务,P(f) 仅包含所示行和第 f 行所代表的索引,代表同一原始样本的不同视图。针对各种应用场景,任务特定的目标 Lts 由不同的优化函数实现。例如,我们在应用MPK-GNN进行分类任务时,采用流行的交叉熵作为优化目标,Lts的形式如下所示:
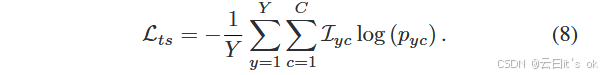
这里,Y 表示训练样本的数量,C 代表类别数量,pyc 是节点 vy 属于第 c 类的预测概率。此外,I 是一个指示函数,如果 vy 的标签是 c,则 Iyc = 1。最后,所提出的多组学学习框架的综合优化损失可以表述如下:
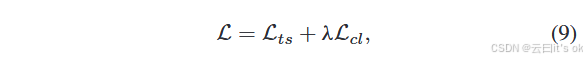
其中λ是一个系数,用于调整对比学习目标的重要性。MPK-GNN的权重可以通过反向传播过程更新。因此,通过优化综合目标,学习到的表示更适合目标任务。
实验
本节通过癌症分子亚型分类任务及其在乳腺癌中的应用,展示了所提出的MPK-GNN框架的有效性。我们首先介绍实验设置,然后展示了所有算法的分类结果,证明了MPK-GNN的优越性。最后,我们还进行了若干额外实验,分析不同超参数或实现的灵敏度。
实验配置
数据集:广受欢迎的TCGA泛癌数据集29,包含基因表达和CNV数据在内的两种组学,用于解决癌症分子亚型分类任务。我们紧密遵循Li等人15所用的预处理步骤。基因表达数据包含11,060个样本,CNV数据包含10,845个样本。在筛选缺失数据后,最终使用的数据集共包含9,664个样本。所有样本包含17,944个遗传特征,属于28个类别。然后,根据它们之间的方差,在对使用的多组数据进行归一化后,选择了最主要的m个遗传特征。具体来说,特征m的数量以{100、200、500、1,000、2,000、5,000}为单位变化,对应的数据集记为TCGA-m。在识别乳腺癌亚型方面,乳腺浸润性癌(BRCA)数据集包含606个患者样本,实验中使用了两种组学(mRNA和CNV)。BRCA包含五种乳腺癌亚型,分别是腔内A型、腔内B型、HER2(+)、TNBC和不明型。mRNA和CNA均由58个遗传特征代表。实验中利用了三种先前的生物学知识,包括基因-基因相互作用(GGI)网络、蛋白质-蛋白质相互作用(PPI)网络和共表达(CoExp)网络。GGI图源自BioGrid数据库26,PPI图和CoExp图均由STRING数据库构建27。
比较方法:为验证所提MPK-GNN框架的有效性,我们比较了两种分类任务在不同方法下的表现,方法涵盖传统机器学习方法到最先进的深度学习模型。具体来说,SVM、射频和KNN都是经典的单视角学习方法。相反,AMGL、MVAR、MLAN、Co-GCN和ERL-MVSC属于多视图学习算法,被提出用于图像或文本分析。最后,DeepMO、MOGONET和CMSC是近期提出的多组学数据分析方法。
实现细节:对于单视图基线,输入的多组学数据将被串接成统一的特征矩阵。为实现公平对比,我们对所用对比方法进行了超参数调优,除了无参数方法AMGL外。例如,射频中使用的决策树数量被调为20棵,而最近邻树的数量被定为1,适用于KNN。请注意,调优实验是在TCGA-1000上进行的,训练样本占10%,然后调优后的数值会应用到所有不同标注比例的数据集上。对于所提方法,基于样本的学习模块的隐藏维数为1,024,所有数据集的256。在特征级学习模块中,图卷积编码器的维分布分别为2048和512,最大池化层的大小设置为8。扁平层的隐藏维数设为32,共享投影函数有两个层,大小分别为2048和32。在训练过程中,SGD优化器的动量设置为0.9,微批量规模调为128。与比较方法的设置类似,上述MPK-GNN的超参数在TCGA-1000上以10%的训练样本调优,然后用于其他情境。我们使用了流行的深度学习软件PyTorch(1.8版本)来实现该方法。对于所有算法,我们每个实验重复十次,并以平均值和标准差结果作为最终表现。所有实验均在Ubuntu-16.04系统上进行,该系统配备2.1 GHz Intel Xeon处理器,配备128 GB内存,以及配备16 GB内存的Nvidia Tesla P100 GPU。MPK-GNN 的实现可在 https://github.com/Xiaoshunxin/ MPK-GNN 获取。
癌症分子亚型分类
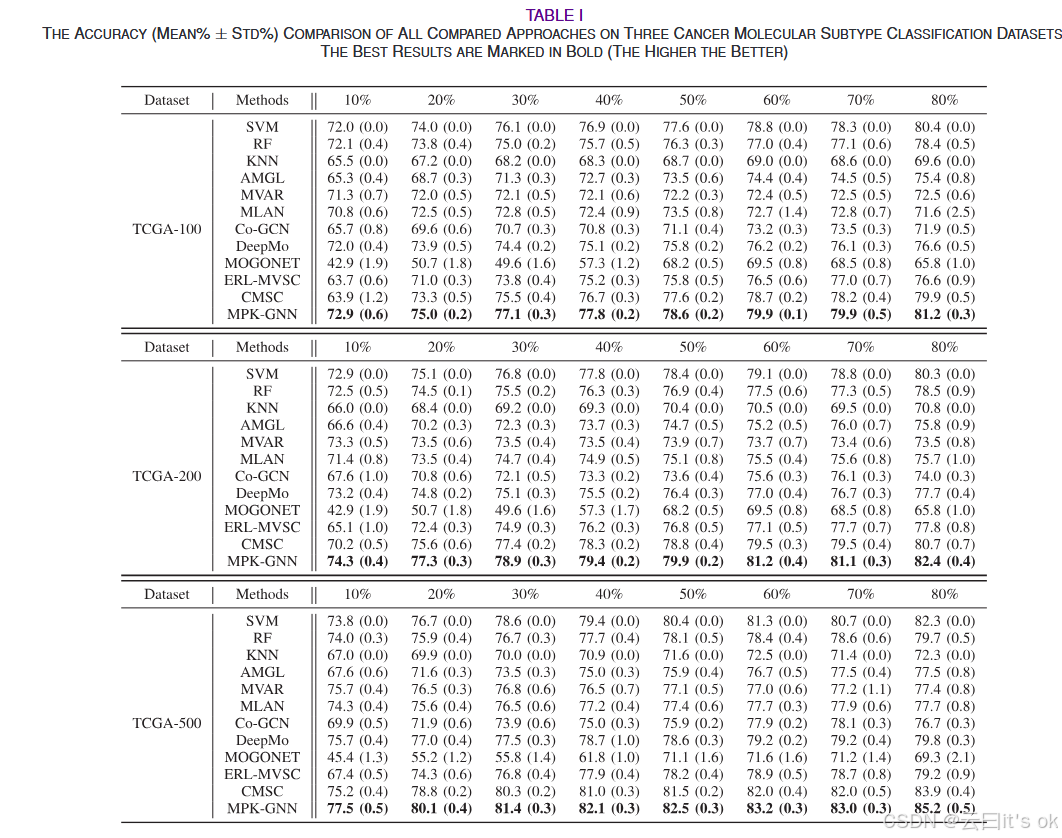
进行了全面实验以验证所提MPK-GNN方法在癌症分子亚型分类中的表现。在表I和表II中,六个数据集上所有比较算法的分类结果分别以不同百分比的训练样本报告。为了直观地比较,最佳结果以加粗标示。MPK-GNN在不同标注率下,在所用数据集中取得了最佳效果。它证明了MPK-GNN能够在半监督和监督任务中取得显著性能。此外,通过分析分类结果,可以观察到一些有趣的现象。首先,所提出的多组学学习框架在所有比较单视角方法中表现优于所有比较。然而,并非所有多视图或多组学方法在不同标记率下都能优于单视图基线的性能。这表明正确利用输入多特征对于多组学数据分析至关重要。与其他基于深度学习的方法相比,MPK-GNN在多次运行下实现了最小的平均标准差。在这方面,所提框架对这些方法更具韧性。需要注意的是,深度学习框架的稳健性对于生物信息处理的应用非常重要。此外,CMSC在TCGA-100和TCGA-200的低标注率(如10%和20%)下表现较差。一个可能的原因是缺乏足够的监督信息来训练学习模型。具体来说,上述环境中训练样本数量和基因特征数量都较少。然而,所提的MPK-GNN在相同情况下通过引入更多先验信息实现了显著性能。这表明,利用多项先验知识对于多组学数据分析至关重要,尤其是在半监督学习的情况下。作为另一种基于GNN的方法,MOGONET在所有情况下的性能都不如CMSC和拟议的MPK-GNN。一个可能的原因是,MOGONET中用于消息传递的邻接矩阵是由节点对之间的余弦相似度构造的。然而,这些模拟的邻接图容易受到输入特征空间质量的影响,可能导致信息传播不正确。另一个可能原因是MOGONET未能利用现有的先验信息来指导学习过程。这进一步表明了多重先验知识的优越性。与使用一个图相比,使用多个先验知识图的好处并不显著,尤其是在遗传特征或训练样本数量较多时。一种可能的解释是,随着训练样本或基因数量的增加,模型接收到的信息量增加,从而降低使用多张先前图的有效性。
乳腺癌中的应用
为验证所提出的多组学数据分析模型的优越性,我们进一步比较了多种学习方法在乳腺癌亚型分类任务中的表现。在表III中,我们报告了BRCA数据集上11个不同标记数据比例算法的分类准确性。实验结果表明,所提出的MPK-GNN框架在不同标记率下优于对比方法。该观察表明,MPK-GNN在特定应用中也有效,如乳腺癌分析。此外,CMSC在大多数情况下表现优于其他比较方法。一个可能的原因是CMSC也将先验知识图谱作为输入。这再次证明了利用先验知识网络的重要性,尤其是在遗传特征或训练样本数量较少的情况下。
模型分析
本节设计了若干额外实验以验证所提出的多组学学习框架。注意,大多数实验环境与IV-A部分的陈述相同,唯独当前分析考虑的因素有所差异。
多项先验知识的影响:我们进行额外研究以验证同时使用多项先验知识的有效性。具体来说,CMSC方法用于获得仅在学习框架中使用一项先验知识时的基线性能。这里,CMSC仅以三项先验知识为参考,包括GGI、PPI和CoExp。此外,我们用常用的GCN替换了CMSC之前基于图的模块,以释放对先验知识的要求。我们将基线记为Basic,其中所需的邻接矩阵是基于输入特征空间构建的。如表IV所示,MPK-GNN在所有基准数据集上,在不同标注比率下均优于所有单一先验知识变体和基线。与单一变异相比,当基因特征和监督样本信息都很少时,所提的多先验知识方法取得了显著提升。通过使用CoExp和GGI网络作为输入图,模型通常比使用PPI网络的模型具有更好的预测准确率。这一现象可以归因于PPI图内的相互作用仅涉及与蛋白质相关且具有已知相互作用的基因,而缺乏蛋白质关联和相互作用的非编码基因则被忽略。此外,我们可以观察到,该方法在所有情况下都优于基线,而单一先验知识则无法达到相同的效果。这一观察表明利用多项先验知识的重要性。
对比学习的影响:为验证共享对比学习框架的有效性,我们开展一项研究,比较所提出的模型MPK-GNN模型的表现,其变体缺乏对比学习损失。具体来说,这种变体丢弃了投影模块,每个输入的先验知识图谱都有自己的学习网络。图2展示了MPK-GNN与该变体的性能比较,实验中该变体表示为无。实验结果表明,所使用的对比学习框架在大多数情况下可以提升学习算法的性能。它表明,对比学习可以通过最大化先有网络之间的一致性,更好地嵌入多个先验知识图中的信息。与无版本相比,设计的MPK-GNN模型网络参数更少,因为对比学习框架被所有先前知识图谱共享。
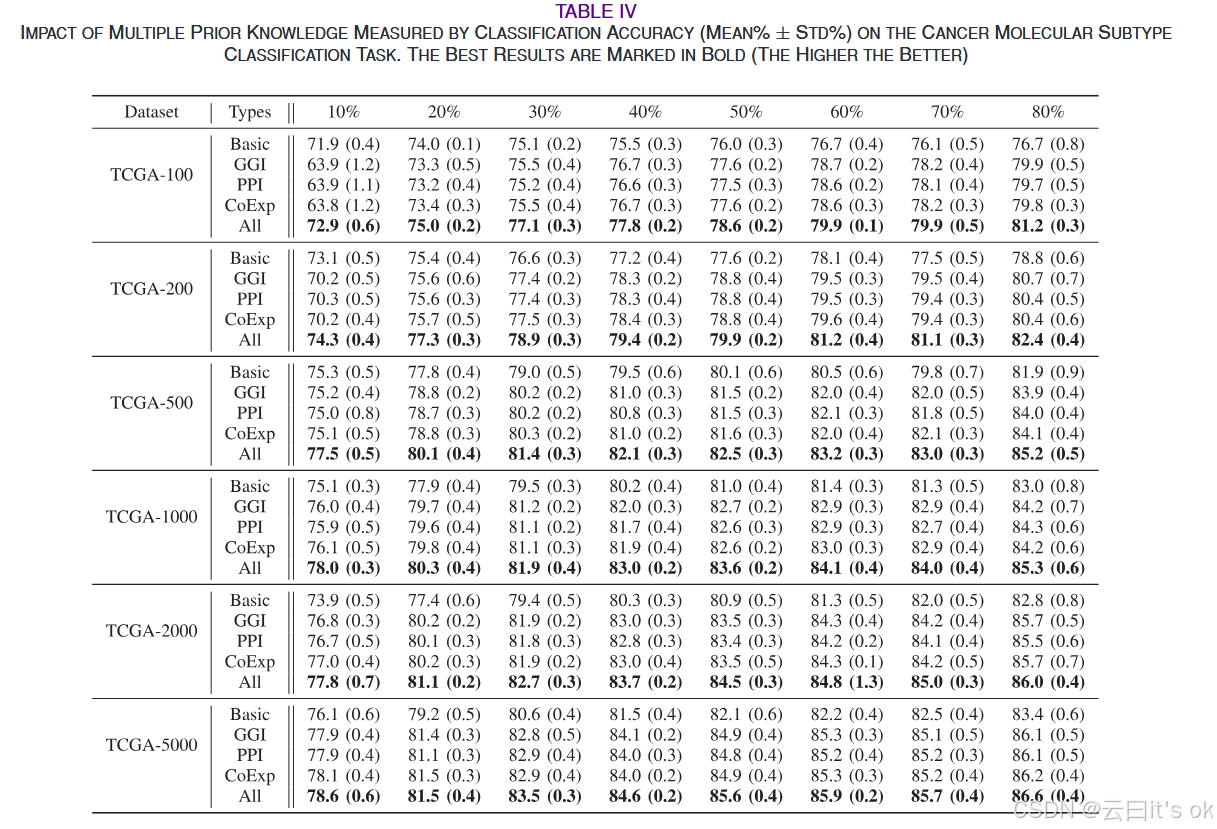
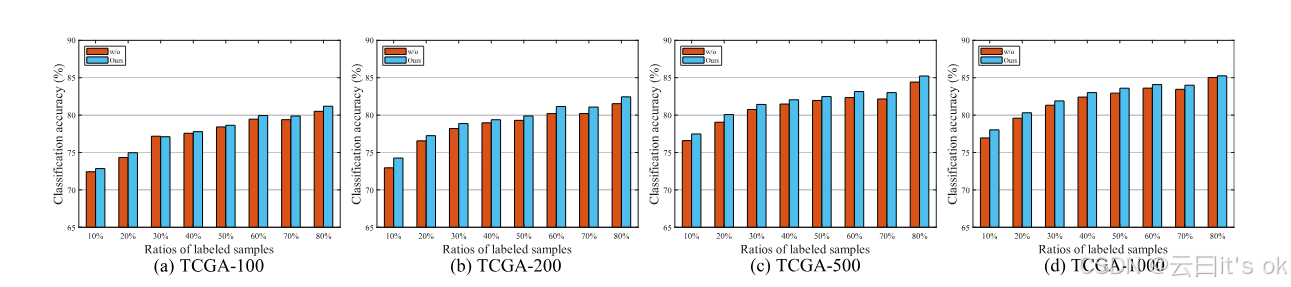
图2。MPK-GNN及其变体在不同标记比率下无对比学习的分类准确性比较
预测标签分布分析:我们进行额外实验,通过分析预测标签分布,研究所有类别的错误分类。本研究基于TCGA-1000数据集进行,80%的训练样本和10%的样本用于测试。基于测试集,我们使用真实类和预测类生成混淆矩阵,该矩阵针对每个类别标准化。如图3所示,五个最容易误入歧途的类别用深蓝色框标出,包括C7:混合(Chr 9 del)、C1:STAD(EBV-CIMP)、C17:BRCA Chr 8q amp、C21:DLBC和C13:混合(Chr 8 del)。其中两种是混合分子亚型,包含多种传统起源癌细胞类型。这一观察表明,预测混合癌症类别而非其他亚型更具挑战性。
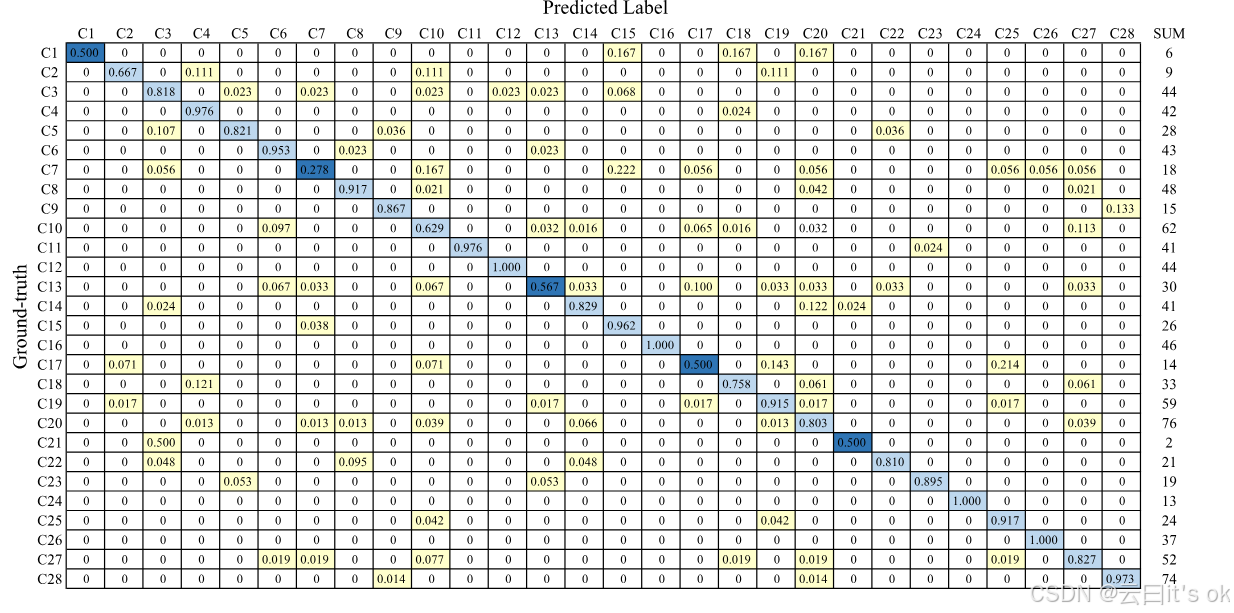
图3。行归一化混淆矩阵,源自TCGA-1000测试集,训练样本为80%。
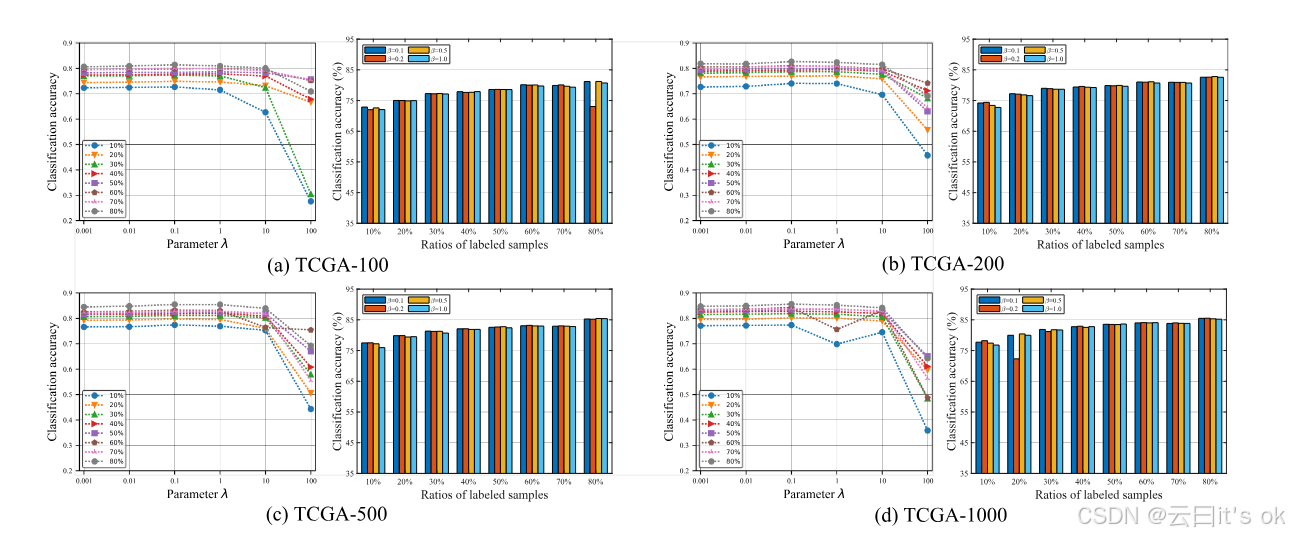
图4。通过不同标签比例下的分类准确性测量MPK-GNN参数λ和β的敏感性分析。
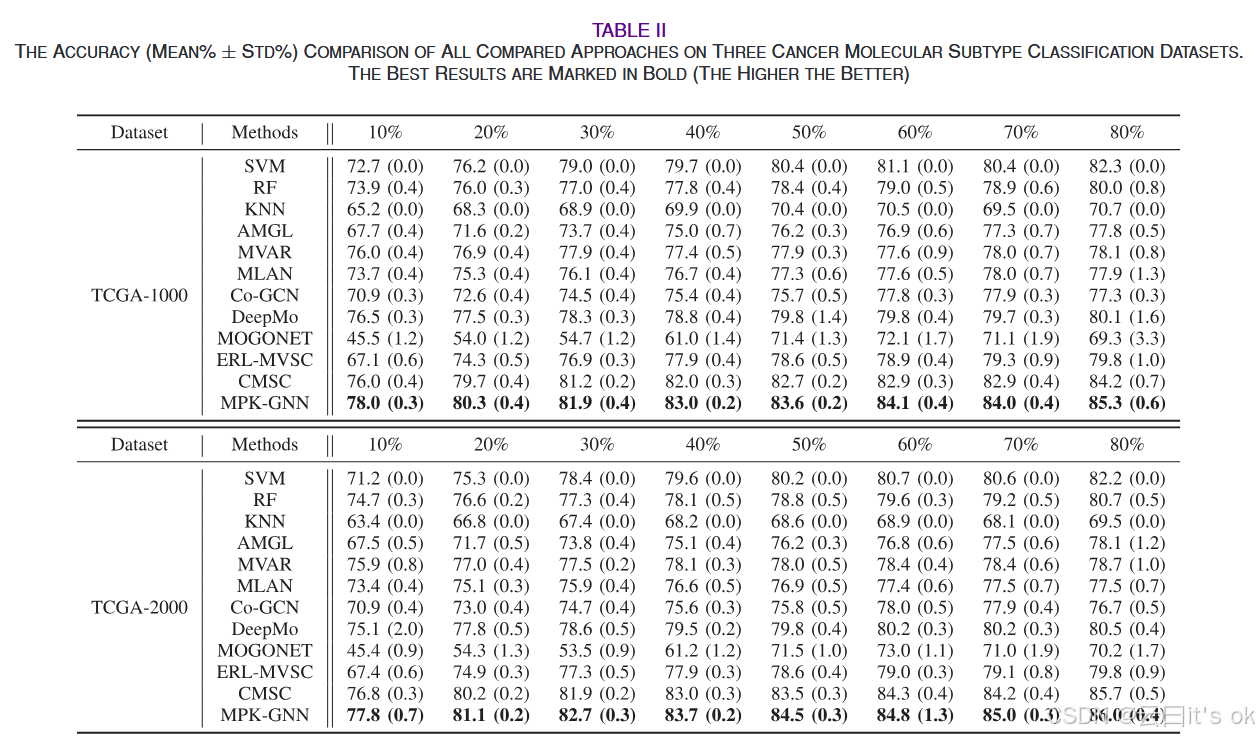
超参数λ和β的敏感性:为验证对对比学习损失λ权重和温度参数β的分类性能敏感性,我们在包括TCGA-100、TCGA-200、TCGA-500和TCGA-1000在内的四个实验数据集上运行了MPK-GNN框架,采用不同组合。具体来说,λ的取值范围为0.001到100,超参数β范围为{0.1, 0.2, 0.5, 1.0}。我们在分析另一个超参数的灵敏度时,固定一个超参数的值。图4展示了直观观察结果。我们可以发现,在大多数情况下,所提出的MPK-GNN框架在改变λ和β值时表现稳定。这一特性对于基于深度学习的多组学分析算法非常重要,因为它们容易受到超参数值的影响。在这方面,MPK-GNN的鲁棒性使其更适合扩展到不同的多组学数据分析任务。 分类性能的统计检验:此外,我们应用Bonferroni-Dunn事后检验,验证癌症分子亚型分类任务中所有算法之间的显著差异。具体来说,如果算法的平均排名距离超过关键差异(CD),则两者性能存在显著差异。CD值由CD = qα √a(a + 1)/6 N计算,其中qα为临界值,a表示比较算法的数量,N表示数据集数量。详细来说,我们比较了除单视图方法外的所有算法,共计九个算法。不同标注比率不同的数据集结果都被用于比较,共计48个数据集。根据30中的表5,当 α = 0.1时,CD = 1.3964。比较结果见图5,横轴刻度值代表平均排名。我们可以观察到,从统计角度看,该算法的性能显著优于除CMSC外的所有比较算法。
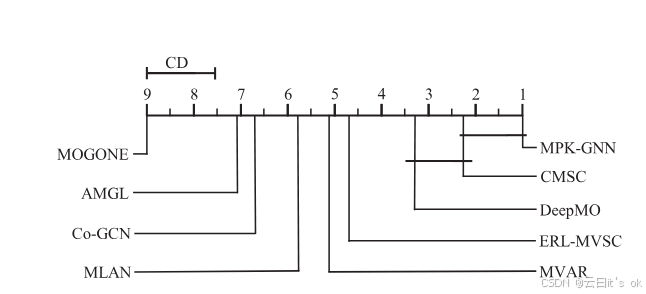
图5。使用Bonferroni-Dunn检验对不同学习算法分类准确度的统计比较。
结论
本文提出了一个基于GNN的深度学习框架,称为MPK-GNN,用于多组学数据分析。设计的共享对比学习架构高效地捕捉了多种先前生物学知识的有用信息,以提升计算模型的性能,尤其是在有少量特征或训练样本可用时。基于多层次表示学习模块,所提出的MPK-GNN框架可以通过配合合适的网络实现任务专属性,灵活地扩展到其他多组学分析任务。综合实验表明,所提框架在七个基准数据集上同时取得了竞争性成果。我们未来的工作包括完善基于样本的模块,以捕捉更多输入多特征的信息,并验证MPK-GNN在更多多组学数据分析任务中的优越性