《智能的理论》全书转至目录****
不同AGI的研究路线对比简化版:《AGI(具身智能)路线对比》,欢迎各位参与讨论、批评或建议。
双手是人类完成动作的主要器官,我们每天的活动都离不开手部活动,如刷牙、梳头、使用勺子搅拌等。刚出生的婴儿就会下意识地摸自己的脸,几个月的婴儿就会有意识地去够东西和抓东西。Jeannerod(1997)作为手部动作研究的先驱者,提出了够物(伸手抓取物体)可以分解为够取(手的传送动作)和抓握(手把物体抓住)两个成分。够取阶段主要依赖于物体的位置信息,而抓取需要根据物体的大小、形状、材质等信息。
一.一个够取和抓握的例子(Shumway-Cook 和Woollacott(著),刘浩等人(译),2021)
通过一个例子说明个体处理够取和抓握的大致流程。例如,当你渴了,想从你面前的牛奶盒中倒一些牛奶到杯子里。
来自周围的感觉输入告诉你周围发生了什么,你所处的空间位置,以及你的关节彼此间的位置关系。视觉系统中有两条平行的通路,一条是与感知和物体识别有关的腹侧通路,另一条是与物体空间定位以及与手部操作相关的背侧通路。感知和识别的通路从视觉皮质向颞叶皮质传导,而定位和动作的通路从视觉皮质向顶叶传导。皮质中的更高级中枢接收到这些信息(可能是顶叶和前运动皮质)并且制订出与目标相关的动作计划:够取和抓握奶盒。这个计划被传到运动皮质。同时它们也会被传到小脑以及基底神经节,它们来调整运动计划使运动更加精细。小脑将调整后的运动输出信号传送至运动皮质和脑干。随后,运动皮质和脑干的下行通路激活脊髓网络,脊髓运动神经元激活特定肌肉,然后你伸手去够取和抓握拿牛奶盒。当你认为牛奶盒是空的但其实很满时,脊髓反射通路会补偿那部分你没有想到的重量,激活更多的运动神经元。然后,你的抓握到物体得到的感觉信号被反馈至小脑,小脑将修正运动输出以适应"变重"的牛奶盒。
二.定位目标(Shumway-Cook 和Woollacott(著),刘浩等人(译),2021)
1.眼-头-身协调
为了能成功够取物体,我们首先必须在空间中定位这个物体。通常,视觉用于物体的定位并引导手的移动(以够物、抓握和徒手操控)。当目标在我们的中心视野时,目标定位通常只涉及眼的运动;而当够取目标位于我们的外周视野时,眼和头都要运动;当目标距中央视野更远时,需要眼、头和躯干的联合运动。
运动学研究显示,当物体出现在外周视野时,由于眼球运动的潜伏期最短,所以它的运动先于头部运动。并且眼睛移动得非常快,所以眼睛首先移动到物体上,并在头运动结束之前,眼睛会一直注视着目标(Jeannerod,1990)。有证据显示,颈部肌肉通常要比控制眼球活动的肌群早激活20~40毫秒。然而,由于眼球的惯性比头部小,所以即使颈部肌群潜伏期更短,仍然是眼球先运动。另外,当需要头部运动来观察一个物体时,头运动的幅度通常只有距目标物距离的60%~75%(Biguer,Prablanc和Jeannerod,1984;Gresty,1974)。
2.参照系
大脑需要做怎样的运算才能做到准确的够物?首先,它需要确定手的位置和目标物(图中门把手)的位置。如图1所示(Buneo和Andersen,2006),这两个位置分别可以用眼睛和身躯两个参照系定义。"T"表示目标的位置,"H"表示手的位置,那么在以眼睛为中心的参照系"E"中,目标的定位可以表示为"TE",手的定位表示为"HE";在以身体(图中的右肩关节)为中心的参照系"B"中,"TB"表示目标位置,手的定位表示为"HB"。手的够取位移"M"即可为"TE-HE",也可为"TB-HB"。
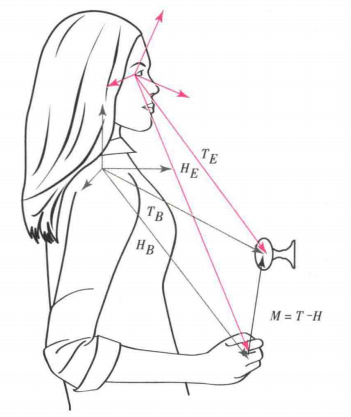
图1
研究表明,后顶叶皮质中的一些神经元编码以眼睛为中心的参照系,该参照系定位手和目标位置;还有一些后顶叶皮质神经元编码以肢体为中心的参照系,该参照系与够物有关的信息。其他后顶叶皮神经元在两个参照系之间的空间信息转换方面起重要作用,协调视觉定位与够物信息(Buneo和 Andersen,2006)。神经系统在够物的早期过程中通过使用以眼睛为中心的参照系作为常规的坐标系统,这是因为视觉是所有感觉中定位最准确的,并能提高运动精确度(Andersen和 Buneo,2002)。另外,有证据表明至少在某些情况下,可以使用眼睛定位的坐标体系,将目标位置"TE"减去手部位置"HE"得到"ME",然后并创建以肢体为中心参照系的运动矢量"MB"。
三.两种操作动作表征(於文苑,刘烨和傅小兰,2018)
1.知觉-动作双分离
视觉系统中存在两个功能不同的亚系统,一个是从枕叶(视觉皮质)投射到颞叶的腹侧通路,该系统主要与客体识别有关,表征物体的视觉特征;另外一个是从枕叶(视觉皮质)投射到顶叶的背侧通路,主要与客体的空间定位有关,表征与物体有关的操作动作信息。这种参与有关物体操作动作信息的表征也被称为操作动作表征。在腹侧通路中,有信息会输人到背侧通路中,与背侧通路中的空间信息和动作信息结合,在背侧通路中形成物体的操作动作表征(Kristensen,Garcea,Mahon和Almeida,2016;Mahon,Kumar和Almeida,2013)。相反,操作动作表征也是物体表征的一个关键部分,对物体识别具有重要作用(Cloutman,2013),表现为被动观看物体手部操作动作时对目标物体的识别有促进作用。
腹侧通路的视知觉表征与背侧通路的动作表征是双分离的。Coodale和Milner(1992)提出了知觉-动作模型。根据此模型,视知觉和视觉引导的动作受视错觉影响的程度应该是不同的,视知觉受影响较大,而视觉引导的动作受影响较小或不受影响。Agioti(1995)首先用铁钦钠圈错觉对该模型进行了验证。在传统的铁钦钠圈错觉中,当两个目标圈的实际直径一样大时(目标圈是被大圆圈或小圆圈所围绕的中间的圆圈),相比被大圈围绕的目标圈,被试的知觉会判断被小圈环绕的目标圈的直径更大,如图2。在Agioti的实验中,用三维的塑料圆盘(可握抓)取代了目标圈。实验要求被试判断两个目标盘哪个大?如果同样大就伸手抓握左边一个;反之抓握右边一个。结果当两个目标盘的实际直径一样大时,被试知觉上认为被小圆圈围绕的目标盘更大,而当被大圈围绕的目标盘的实际直径稍大时,被试则倾向于判断两者相等。尽管知觉上两目标盘的直径受错觉影响,但是在被试伸手抓握过程中的最大抓握直径均接近于目标盘的实际大小。这说明,相比视知觉,动作表征受视觉错觉影响较少,支持知觉-动作模型。

图2
2.腹侧通路与背侧通路的配合
为了完成一次够取动作,需要腹侧通路与背侧通路的共同配合。可以通过在咖啡厅拿起咖啡时的活动了解这个过程。假设小明需要拿起一杯咖啡。如图3,首先,他从桌子上的花盆和其他物体中识别出咖啡杯(腹侧通路)。一旦咖啡杯被感知到,他就需要思考它在桌子上的位置并伸手碰触它(背侧通路)。当他避开花、拿咖啡杯,确定好手和手指的方位去抓住杯子(背侧通路)时,他需要对杯子的手柄进行知觉加工(腹侧通路)。接着,他会根据杯子里咖啡的多少(腹侧通路)施以恰当的力量(背侧路径)将杯子拿起来。(Goodale,2011)。

图3
3.背侧-背侧通路和腹侧-背侧通路
有研究表明,视觉的背侧通路可进一步分为背侧-背侧通路和腹侧-背侧通路,前者利用当前物体的视觉和空间信息,形成物体的结构性操作,这种操作包括对物体的够取和抓握。该通路又被称为抓握系统;后者利用物体的使用经验和功能性特征(如按键是拿来按的),实现人对物体的功能性操作,即使用物体以发挥其特定的功能(Brandi,Wohlschläger,Sorg和Hermsdörfer,2014)。该通路又被称为使用系统。基于背侧通路的这两条子通路,操作动作表征可以分为结构性操作动作表征和功能性操作动作表征(Binkofski和Buxbaum,2013;Buxbaum和Kalénine,2010)。
4.结构性操作动作表征
结构性操作动作系统通过处理当前输人的物体视觉和空间信息,包括物体的形状、大小、位置,以及物体与视网膜和躯体的相对位置,形成与物体相符的够取和抓握动作的表征,即结构性操作动作表征。结构性操作动作系统仅依赖当前的信息就可以形成结构性操作动作表征,对物体进行实时加工,其加工过程不会占用过多的工作记忆资源。另外,结构性操作动作表征可以在无注意条件,甚至可能在无意识的条件下被激活,但是激活后维持的时间比较短。
5.功能性操作动作表征
功能性操作动作系统主要处理和储存与物体相关的使用动作信息。当人通过某一个特定的动作将某一物体使用多次后,功能性操作动作系统会将这个使用动作的核心特征提取并储存起来,形成物体的功能性操作动作表征。功能性操作动作系统需要从长时记忆和概念系统中提取信息,因此其加工过程中需要占用更多的工作记忆资源。该表征在有注意和意识参与的条件下才可以激活,并且在激活后可以维持较长时间。结构性操作动作表征系统与长时记忆系统的联系程度相对较低,但是结构性操作动作表征也可以储存在长时记忆中。而功能性操作动作表征则只能存储于长时记忆中。
6.两种表征与长时记忆关系的实证
面对一个操作冲突物体(结构性操作动作和功能性操作动作不一致的物体),如果仅要求被试摆出抓握动作(结构性)或者摆出使用该物体动作(功能性),被试摆出抓握动作的反应时比摆出使用动作的反应时更短(Jax和Buxbaum,2010);但是如果要求被试真实地抓握并移动物体或者真实地使用物体时,被试真实地做出使用动作的反应时与摆出使用动作的反应时没有显著差异,但是被试真实地做出抓握动作的速度比真实地做出使用动作的速度更慢,也比摆出抓握动作时的速度慢(Osiurak,Roche,Ramone和Chainay,2013)。
这一方面说明,结构性操作动作表征只在某些情况下被提取,在仅仅摆出抓握的动作时,被试利用当前输人的信息就可以形成这个物体的结构性操作动作表征,因此被试摆出该动作的速度较快;而在真实地将物体抓握并拿给他人时,被试还要依赖其他信息(如物体本身的特征信息(如重量)),这就要求被试提取长时记忆中的物体特征和相应的结构性操作动作表征,所以被试做出相应的抓握动作的速度较慢。另一方面,功能性操作动作表征不能仅根据当前输入的信息而形成,必须从长时记忆中提取后才可以使用,所以被试无论是摆出使用动作或是真实地使用物体,都需要较长的反应时(Osiurak,Roche,Ramone和Chainay,2013)。
7.不同信息模态下两种操作动作表征的激活
操作动作表征能被多种模态的信息激活,包括视觉模态、语义模态和动作模态信息(Rey,Roche,Versace和Chainay,2015)。而两种操作动作表征对于不同模态的信息会分别表现出不同的激活状态。首先,被动观看物体图片可以激活结构性操作动作表征,说明结构性操作动作表征能被单独的视觉模态信息激活(Mcnair和Harris,2012),而被动观看物体图片不足以激活功能性操作动作表征,需要被试进行一定的操作动作才可以激活,说明功能性操作动作表征在视觉模态和动作模态信息结合的条件下才可以激活(Squires,Macdonald. Culham和Snow,2016)。其次,被动观看物体名称不能激活两种操作动作表征,但被动观看句子中的物体名称可以激活功能性操作动作表征。经过语义判断的物体名称也可以激活两种操作动作表征(Bubet al..2008)。第三,两种操作动作表征都可以分别被结构性或功能性操作动作的手势激活(Bub,Masson和Lin,2015),也能被句子中表达结构性或功能性操作动作的动词激活,说明动作模态的信息能激活两种操作动作表征(Bub和Masson,2010)。
这些结果说明结构性和功能性操作动作表征都能被视觉模态信息激活,但是由于结构性操作动作表征是实时加工,因此其激活更依赖于当前输人的视觉模态信息;而功能性操作动作表征与长时记忆的联系更强,因此它更容易被语义模态的信息激活。此外动作模态的信息会引起更强的运动系统的激活,从而使两种操作动作表征的激活表现得更加明显。
四.够取(Shumway-Cook 和Woollacott(著),刘浩等人(译),2021)
当视觉对手和目标进行定位后,会计算出手部够取的动作。距离编程理论和定位编程理论是解释手部够取的两种理论。
1.距离编程理论
根据距离编程理论,当手臂去够取目标物时,人们在视觉上可感受到要移动的位移,并激活一组特定的主动肌以驱动手臂到达距目标物一定的范围内。随后,他们会放松主动肌并激活对应的拮抗肌以提供停止运动的制动力(Keele,1986)。总之,距离编程理论认为,编码够取的神经编码是关于手的位移信息的,该位移信息指导手的够取位置。在距离编程理论的框架下,包括多重纠正理论、脉冲可变模型和混合模型等子模型。
(1)多重纠正理论
多重纠正理论强调,在移动过程中,视觉反馈对于动作纠正和够取准确率提升的重要性。因此,不准确的运动可能是视觉反馈缺失的结果。
当视觉缺失时,手臂运动的准确性会降低。对于四肢患有严重的外周感觉神经病变的患者,当要求他们闭着眼睛时,患者也能完成各种各样的简单动作,如敲打和用手指在空中画画(开环控制)。但是,当要求他们闭着眼睛多次重复某个动作时,他们的表现会迅速变差(需要闭环控制)。这不但提示我们,只要动作是简单的或不需要重复的,躯体感觉信息便不参与上肢运动的执行;同时也提示我们,如果患者要完成由多个关节协调运动的动作或重复某一动作,没有视觉反馈他们将无法辨别躯体空间感觉的表征是否符合目标动作表征,并且会表现出大量的动作"偏航"以及协调问题(Rothwell,Traub,Day和Obeso等人,1982)。此外,当要求受试者进行不同持续时间的手臂运动时,190亳秒或更短时间的手臂运动不会因为视力缺失而受到影响,而260毫秒或更长时间的运动会受视觉反馈缺失的影响(Keele和Posner,1968)。
因此,运动轨迹的修正是建立在视觉反馈的基础上的,并且需要200~250毫秒的时间来不断更新运动轨迹。在20世纪60年代,研究人员(Keele,1968)提出,瞄准运动(包括够取)由一系列子运动组成。第一个子运动(进行任何视觉校正之前)的位移覆盖到至目标的大部分距离,并且与最终的精确度无关。但后续每个子运动都可减少视觉误差(第二次子运动纠正第一次子运动的误差,第三次纠正第二次的误差...),最终保证瞄准运动的顺利完成。由于瞄准运动的时间通常很短,因此通常情况下瞄准目标的运动只有1次校正(如果有的话)(Rosenbaum,1991)。Woodworth(1970)的两阶段模型认为,瞄准运动由初始脉冲阶段和视觉反馈控制阶段组成。他认为最初的脉冲阶段在中枢神经的控制下,被编码成肢体接近目标的信息。一旦进入目标的附近区域,肢体就会受到基于反馈的视觉信息的控制。在第二个反馈阶段,利用肢体和目标相对位置的视觉信息对运动轨迹进行调整,使肢体准确的到达目标。这些调整可以采取"小的额外动作"的形式,即其位移距离比第一阶段的更短。
(2)脉冲可变性模型
Schmidt等人(Schmidt,Zelaznik,Hawkins和Frank等人1979)遵循Woodworth的两阶段模型,在初始脉冲阶段是一个不基于反馈纠正过程的模型,该模型认为运动开始前,中枢命令预先设定了到达目标所必须经过的距离,原动肌与拮抗肌产生爆发力(开环控制)推动肢体达到指定目标区域。他发现,当他要求被试进行快速而准确的运动时,所需的力会很大,很大的力会导致力的变异性增加,而增加的力的变异性会导致运动准确性降低(Schmidt,Young,Swinnen和Shappiro,1989)。该初始运动的特征可以在以下等式中描述:
We= a+ b(D/MT)
其中,We表示运动终点的变异性,D是运动的距离,MT是运动时间,a和b是公式的常数,该等式与Fitts定律(14-2:个体、任务与环境)相似。它表明,在快速无视觉反馈的情况下,速度越快的运动,需要的力量越大而运动的准确性越低。该理论表明,当运动幅度D足够短时,即便运动时间较短MT,如果速度(D/MT)足够慢也能保证运动的准确性,从而保证一次瞄准就能成功(无需第二次子运动的反馈)。但仅凭这一理论不能用于解释瞄准运动,正如上面所看到的,许多运动特别是持续时间超过250毫秒的运动确实需要使用视觉反馈来提高准确性。
(3)混合模型
前面的两种模型是处理运动控制的两个极端:(a)在较慢运动的过程中使用视觉反馈以提高准确性;(b)非常快速的运动不容易利用视觉反馈,因此只能通过初始脉冲的幅度来控制(当运动幅度足够短时,可以在短时间内完成动作)。而混合模型则联合这两种模式的要素(Meyer,Abrams,Kornblum和Wright等人,1988),这种混合模式又被称为"优化初始脉冲模式"。
混合模型认为,在一次瞄准运动中,被者向一个目标发起一次运动,如果成功"命中"(成功够取),便完成了这次瞄准运动(只有一次子运动)。然而,如果没有命中(例如手还没达到目标或手超过目标),那么通过视觉反馈,进行下一次子运动以完成瞄准运动。显然,被试者需要在快速运动和缓慢运动之间找到一种平衡,因为快速运动需要很大的初始力而使运动变得不准确,但它只需一次运动;而缓慢运动能够得到校正从而确保准确性,但它需要多次运动(Rosenbaum,1991)。
2.定位编程理论
根据距离编程理论,大脑是通过控制手在原来位置的基础上位移一段距离来够物(大脑编码的是手的位移)。相反,定位编程理论认为,空间中每一个够取位置都由一组相应肌群(原动肌与拮抗肌)的张力(长度)的神经编码定义。换句话说,如果需要手到达某一空间位置,通过神经编码,控制相应肌群进行不同的伸缩,不同伸缩状态的肌群对应手所够取的不同位置,即特定空间位置与一组特定肌群的收缩放松长度对应。可以使用咖啡馆门口弹簧的例子解释定位编程的模型(Keele,1986)。假设,门口关闭是门口的稳定状态(如图4(a)),此时的弹簧长度是它们的自然长度。当对门口施加一个力时,咖啡馆的门就会发生运动,此时一个弹簧的长度减少而另一个弹簧的长度增加,弹簧由自然长度变为非自然长度从而产生弹力。当放开门时,弹簧的弹力使门返回到关闭的位置。如果想让门一直开着(或者说,门开着是它的稳定状态),可以简单地将两个弹簧换成长度不同的弹簧,那么门将处于一个新的稳定位置(如图(b))。关节上成对的原动肌和拮抗肌就像咖啡馆门上的弹簧。我们可以通过不同的相对激活程度来改变这两块肌肉的相对长度,从而改变关节不同屈伸程度的"稳定状态"。比如,用较高激活程度激活原动肌(收缩,肌肉变短)和用较低激活程度激活拮抗肌(伸展,肌肉变长),此时关节处于屈状态,这是该关节在该神经编码下的稳定状态,如用一外力改变该关节的状态(如使它更伸直或更弯屈),当外力撤开后关节又会回到原来的屈状态。而这个关节状态使手处于某一特定位置,因此一组肌肉的长度(激活状态)对应手的一个特定位置;同理,较低激活程度的原动肌和较高激活程度的拮抗肌使关节处于"伸"的自然状态...。因此,仅仅控制肌肉的相对长度,就能使手处于某个特定位置。
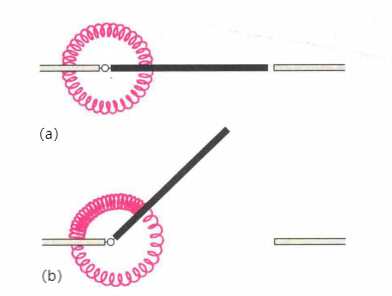
图4
一项关于猴子够取的实验(Polit和Bizzi,1979)中,猴子视线内有若干个灯泡分布在不同位置,研究人员会训练猴子当目标灯泡的灯光时,需要其肘部运动以达到该目标,如图5所示。在正式实验中,在一些实验条件下,猴子戴着阻断手臂视线的颈圈以消除视觉反馈(猴子不能看到自己手的运动)。此外,在一些实验条件下,脊髓后根也被切断,从而阻断手臂的动觉反馈。实验人员会在有和没有视觉或动觉反馈的情况下,测量猴子手臂运动的准确性。从实验结果中发现,尽管失去了视觉和动觉的反馈,它们的够物仍是正常的(如图(b))。
随后,在没有视觉和动觉反馈的条件下,研究人员对猴子的手臂进行干扰,在目标灯亮起后,猴子手臂开始移动之前,人为地将其手臂从原来的位置移开。由于没有视觉和动觉反馈,猴子受到于扰时不能感觉到或看到手臂的位置。因此,如果猴子编码的是距离或位移(距离编程理论),那么猴子在指向目标时手最终的位置应该与目标有一定距离,这个距离大约是手被人为移动的距离。如果编码的是位置,那么它们还是能以适当的准确度够取到目标(如图(c)和图(d))。因为在这种情况下,神经系统编程的是手臂主动肌和拮抗肌的长度,而这组肌群的特定长度对应一个特定位置。一旦形成新的"弹簧装置"(编码的肌群长度),那么不论肢体在哪儿受到干扰,就像咖啡馆的门一样,肢体将会摆动到其神经编码长度。因此,猴子不需要知道它的手的起始位置(被人为移动)便可以到达正确的终点。该实验结果支持定位编程理论。
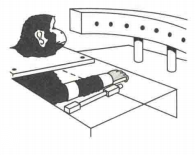
(a)
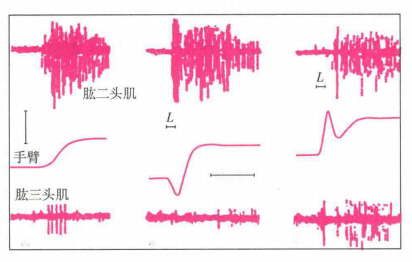
图5(b) 图5(c) 图5(d)
3.联合编程
这些结果是否表明距离编程是错误的?不是的。最可能的是根据任务和环境,距离和定位两种编程策略都用于手臂的运动中。例如已有研究表明,当人类进行快速的肘关节屈曲运动时(Hallett,Shahani和Young,1975),他们表现出三阶段的突发收缩,首先肱二头肌被激活,接着是肱三头肌(制动运动),然后又是肱二头肌。同样的模式在动觉丧失的患者中也出现。然而,当被试被要求移动得更慢和更平稳时,他们表现出连续的肱二头肌活动且没有肱三头肌活动。这使得许多研究人员认为,人们在缓慢的运动中应用定位编程来实现,而更快速的运动则联合应用距离编程和定位编程。
总之,短于0.25秒的单关节运动由于时间太短而无法利用视觉反馈,这种快速移动可能会涉及距离和定位的联合编程;而长于0.25秒的关节运动则在起始阶段会涉及视觉反馈,这种缓慢运动可能涉及定位编程。
五.抓握
1.分类
抓握的模式依据要抓握物体的形状、大小和位置而变化(Johansson,1996)。Napier(1956)将人类的抓握运动分为力量抓握和精确抓握。从解剖结构上讲,两种抓握的不同在于拇指和其余手指的姿势不同。力量抓握时,拇指和手指的指腹朝向手掌,并将力传递到物体上。力量抓握包括钩状抓握(握手提箱的把手)、球状抓握(握住垒球)和柱状抓握(拿着瓶子)。相反,精细抓握时,力量存在于拇指和其余四指之间。这两种抓握在操作技巧中的应用大相径庭:精细抓握可进行物体相对于手的运动,力量抓握却无法完成这些动作。Napier(1956)发现,对于几乎每一类型的物体都可单独使用精确或力量抓握,或者将两者联合使用。他也认为,不只是物体的形状或大小,还有目的性活动本身也决定抓握的类型,如一个柱状物体可以用来写东西(精确抓握)或者用来敲打(力量抓握)。
在猴子完成精细抓握和力量抓握任务的实验中(如图6)已经证实,初级运动皮质区的神经元只在执行精细抓握时才被激活(包括轻力量的精细握抓和重力量的精细握抓),但在力量抓握中则没发现该区域神经元的激活。这说明,这两种握抓有两套不同的神经控制模式。(Muir和Lemon,1983)
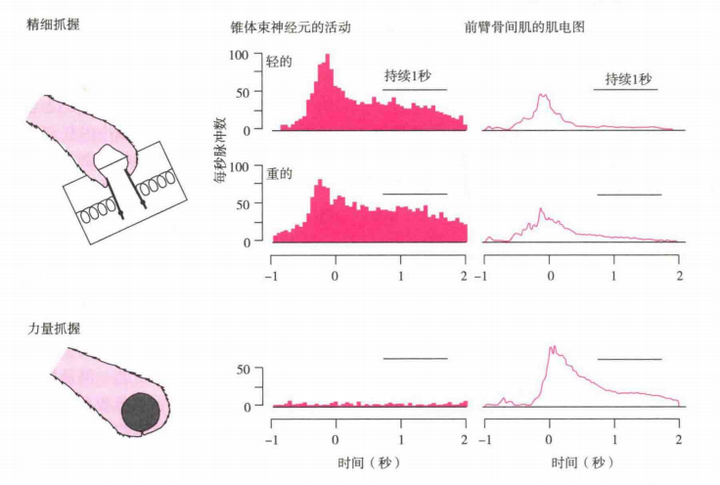
图6
2.抓握的形成过程
当向前去抓握一个物体时,抓握手形的形成发生在够物的移动过程中。在此过程中,双手速度及位移(图7左上)与抓握口径大小改变(图7右上)的关系如图所示。抓握口径大小与目标物体成正比,最大抓握口径与物体的大小是相适应的。例如,图7下分别显示了两个被试在够取直径为2mm的小棒和直径为55mm的圆柱体时,抓握口径的变化。结果显示,不同被试在够取不同直径物体时,抓握口径在够物过程中有相似的变化规律,且物体大小每增加1cm,最大抓握口径随之增加0.77cm。(Marteniuk,Leavitt,MacKenzie和Athenes,1990)。
在够取的过程中,手掌部分拇指的活动程度少于其余四个指头。当受试者改变抓握的口径时,几乎完全靠其余四个手指的运动,而拇指保持不动。随着手接近目标物体,手移动的速度先增加后减少。且抓握口径随着够物过程先增加到最大(大于目标物大小的最大抓握口径),然后减少到适合目标物的大小。这个最大抓握口径一般出现在够取阶段的后半程,并且面对越大的目标物体,最大抓握口径在整个伸出手臂过程中出现的时间点越靠后(Rosenbaum,Cohen,Meulenbroek和Vaughan,2006)。面对同一尺寸的物体,研究人员还发现目标物体的重量也能影响抓握物体的手部运动。去抓握较重的物体时,手掌的最大抓握口径要更大,并且在接触物体时手指更接近物体的重心(Eastough和Edwards,2007)。
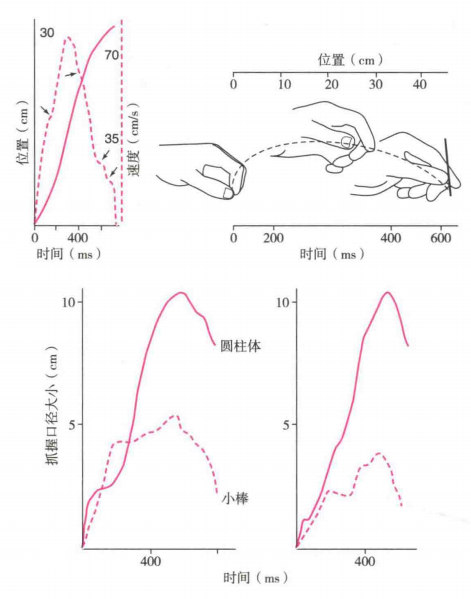
图7
最后,在无法看到物体和手的条件下,被试的最大抓握口径要大于被试可以看到物体和手的条件。这可能是因为在无视觉反馈时,抓取动作的不确定性增大,所以被试有意增大了手指抓握口径以保证抓取的有效性。(唐日新,宋爱霞和陈娟,2019)
六.够取和握抓的协调(Shumway-Cook 和Woollacott(著),刘浩等人(译),2021)
在够物过程中,上肢移动手到达目标物(够取)与抓握物体的手指的预先构形(握抓)是同时进行的。够取和抓握的分离也得到了生理学上的证据支持,比如研究发现大脑顶枕皮层负责够取,而前内顶沟负责抓握。另外,虽然新生儿的抓握动作形成发育较晚,但早期就可以观察到他们的够取动作。研究表明,1周大的婴儿可以够物,并且可以截住移动的物体,开始去触摸它们,但这是在手掌完全张开的情况下做的动作,此时并没有抓握动作形成。抓握动作的形成在婴儿10~22周时才开始发育(Bruner和Koslowski,1972)。虽然够取和抓握是由不同的系统控制,但在整个运动过程中,它们(手的移动与手势形状的形成)彼此间必须相互协调,以确保当手指接触到物体时抓握动作的完成。
成功地抓握一个物体需要满足两个重要的条件。首先,手必须适应物体的形状、大小和用途。其次,手指的运动必须与上肢的移动在时间上有很好地协同,才能在恰当的时刻靠近物体。如果靠近物体的时间过早或过晚,都不适合完成抓握(Jeannerod,1990)。研究发现,最大抓握口径的发生时间相对于整个运动时间有一个固定的比例,它发生在75%~80%的够取运动时间中(Wallace,Weeks和Kelso,1990)。这一比例在不同的运动时间、速度和不同的手指初始姿势中是不变的,即使是在病理条件下也保持不变。这是够取和抓握的功能性联系的有力证明(Jeannerod,1996)。
有研究表明,相比要求指向或击打某一物体,要求被试去抓握一个物体的运动时间要长得多。同样,当准备抓握一个物体时,够取运动的加速时间要比减速时间短,但是如果要求被试击中一个目标时,加速时间要比减速时间长得多,这使得受试者以一个相对快的速度击中目标(Marteniuk,Mackenzie,Jeannerod和Athenes等人,1987)(图8分别显示了够物运动和指向运动的时间速度关系)。此外,如果被试抓握起目标物,然后把它放进一个小盒子里,又或者把它扔掉,扔掉的运动时间比放进盒子的运动时间要短。
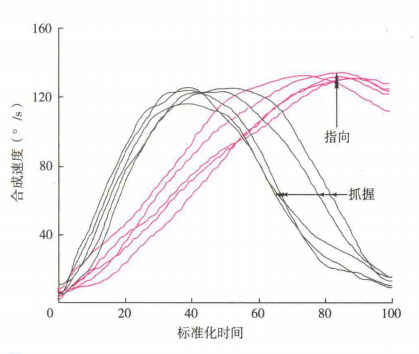
图8
七.拿起(Shumway-Cook 和Woollacott(著),刘浩等人(译),2021)
一天中人们拿起物体的种类繁多,可能是一只很轻的钢笔,也可能是一个又重又滑的油瓶,而神经系统能够精确地调整抓握以适应许多不同重量和表面特性的物体。现已证实,任何一个拿起任务都有四个分离的时相(Johansson和Edin,1992):
(a)第一时相,拿起任务起始于手指和目标物体的接触之前。
(b)第二时相,一旦接触到物体(物体的重量分别为200g、400g和800g),第二时相就开始了,如图9(a)所示,实验中目标物体有抓握力和负荷力(使目标物体向上的负荷)的传感器感知被试手部的力。在该时相中,抓握力和负荷力会逐渐增加,如图(b)所示。
(c)第三时相,当负荷力超过物体的重量且物体开始移动时,第三时相开始,此时物品的位置从0开始向上移动,如图(b)下图所示。然后抓握力和负荷力逐渐下降到适合的大小。
(d)第四时相,此时物体被放下后,抓握力和负荷力迅速下降。
由于不同的物体有不同的光滑度,所以即便是同样重量的物体,所需的抓握力可能也不同。中枢神经系统似乎会结合以往的经验以及拿起物体时的感知觉信息来选择正确的抓握参数(抓握力)。如果预期与实际不相符(如认为物体比预期重或者比预期滑),指腹的感受器就会被激活(物体相对手指有下滑趋势)。由于环层小体非常敏感,因此能够很容易地检测到物体的滑动。通过增加手指肌肉的活动来增加抓握的力量,同时增加肩、肘部的肌肉活动来降低手的加速度,以减少物体的下滑趋势。另外,在将手指麻醉以阻止皮肤感觉反馈的实验中,被试会通过增加抓握的力量以代偿皮肤反馈感觉的缺失(Witney,Wing,Thonnard和Smith,2004)。此外视觉和其他类型的皮肤信号在决定抓握参数的选择时也很重要(Johansson和Edin,1992),如看到物体下滑会加大抓握力等。
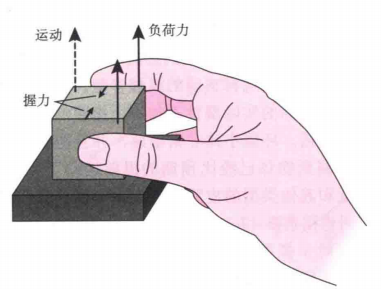
图9(a)
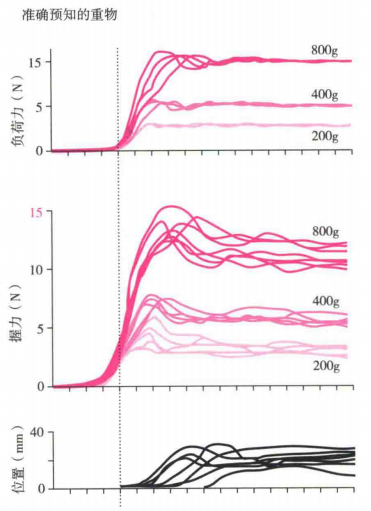
图9(b)